Esperto medico dell'articolo

Nuove pubblicazioni
Modelli sperimentali di osteoartrite
Ultima recensione: 07.07.2025

Tutti i contenuti di iLive sono revisionati o verificati da un punto di vista medico per garantire la massima precisione possibile.
Abbiamo linee guida rigorose in materia di sourcing e colleghiamo solo a siti di media affidabili, istituti di ricerca accademici e, ove possibile, studi rivisti dal punto di vista medico. Nota che i numeri tra parentesi ([1], [2], ecc.) Sono link cliccabili per questi studi.
Se ritieni che uno qualsiasi dei nostri contenuti sia impreciso, scaduto o comunque discutibile, selezionalo e premi Ctrl + Invio.
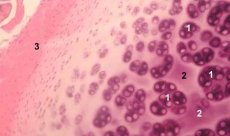
La cartilagine è un tessuto altamente specializzato contenente un solo tipo di cellule (condrociti) e caratterizzato dall'assenza di vasi sanguigni e linfatici. La cartilagine è nutrita principalmente per assorbimento dal liquido sinoviale. Il metabolismo dei condrociti è regolato da una serie di fattori solubili prodotti localmente dai condrociti e dai tessuti circostanti. La funzione dei condrociti dipende anche dalla composizione dell'ambiente extracellulare (tensione dell'ossigeno, concentrazione di ioni, pH, ecc.), dalla composizione della matrice extracellulare (ECM), dall'interazione tra cellule e matrice e dai segnali fisici. L'obiettivo principale della modellizzazione sperimentale è creare colture nell'ambiente extracellulare senza alterare il fenotipo delle cellule mature. Il secondo obiettivo è creare colture per studiare la risposta prematura, ritardata, a breve termine o prolungata dei condrociti a segnali chimici e/o fisici. Gli studi in vitro offrono anche l'opportunità di studiare il comportamento dei condrociti nell'osteoartrosi. Il terzo obiettivo è sviluppare sistemi di co-coltura che consentano di studiare le interazioni tra i vari tessuti nell'articolazione. Il quarto obiettivo è la preparazione degli impianti cartilaginei per il successivo trapianto. Infine, il quinto obiettivo è lo studio di fattori di crescita, citochine o agenti terapeutici in grado di stimolare la riparazione e/o inibire il riassorbimento cartilagineo.
Negli ultimi decenni sono stati creati vari modelli di colture cellulari di cartilagine articolare, tra cui colture monostrato, colture sospese, colture di condroni, espianti, co-colture e colture cellulari immortali. Ogni coltura presenta vantaggi e svantaggi specifici, ed è adatta allo studio di un aspetto specifico del metabolismo dei condrociti. Pertanto, gli espianti di cartilagine rappresentano un modello eccellente per lo studio del turnover degli elementi della matrice, che richiede recettori di superficie cellulare autentici e normali interazioni cellula-matrice e matrice-cellula. Allo stesso tempo, si raccomanda di studiare i depositi di matrice o i meccanismi che regolano il metabolismo dei condrociti su una coltura di cellule isolate. Una coltura monostrato a bassa densità è necessaria per studiare il processo di differenziazione cellulare. Le colture sospese in una matrice naturale o sintetica rappresentano un modello per l'analisi della risposta adattativa dei condrociti allo stress meccanico.
Colture di condrociti
Diversi punti importanti devono essere considerati nella selezione del tessuto cartilagineo per gli studi in vitro. La composizione della matrice e l'attività metabolica dei condrociti variano tra le articolazioni, e quest'ultima dipende anche dalla profondità della localizzazione dei condrociti nel tessuto. Questi dati sono stati ottenuti in diversi esperimenti in cui sono state studiate sottopopolazioni di condrociti isolati da zone cartilaginee di diversa profondità. Sono state riscontrate diverse differenze morfologiche e biochimiche tra i condrociti in coltura situati negli strati superficiali e profondi della cartilagine articolare. Le cellule superficiali sintetizzano una matrice fibrillare rada e povera di proteoglicani, mentre le cellule più profonde producono una matrice ricca di fibrille e proteoglicani. Inoltre, le cellule superficiali producono relativamente più piccoli proteoglicani non aggregati e acido ialuronico e relativamente meno aggrecano e cheratan solfato rispetto ai condrociti più profondi. Un'altra importante caratteristica distintiva del metabolismo dei condrociti isolati da zone cartilaginee di diversa profondità è la risposta a uno stimolo esogeno. Secondo M. Aydelotte et al., i condrociti bovini della zona superficiale della cartilagine erano più sensibili all'IL-1 rispetto alle cellule della zona profonda.
Il comportamento cellulare dipende anche dalla posizione del tessuto. I condrociti della cartilagine costale e auricolare dello stesso animale rispondono in modo diverso a fattori di crescita come il fattore di crescita dei fibroblasti (FGF) e il TGF-β. L'FGF ha aumentato l'incorporazione di timidina, prolina e leucina nelle costole coltivate, ma non nei condrociti auricolari. Il TGF-β ha aumentato l'incorporazione di timidina nei condrociti della cartilagine costale e auricolare, ma non ha avuto effetto sull'incorporazione di timidina e prolina nei condrociti auricolari. Le cellule cartilaginee provenienti da aree ad alto stress differiscono da quelle provenienti da aree a basso stress cartilagineo. Pertanto, i condrociti della cartilagine articolare del ginocchio di pecora matura, provenienti dalla regione centrale della superficie articolare della tibia non ricoperta dal menisco, che sopporta il carico maggiore in vivo, sintetizzano meno aggrecano, ma più decorina rispetto alle cellule provenienti dalle zone rivestite dal menisco. Gli autori sottolineano inoltre l'importanza di utilizzare cartilagine proveniente da zone articolari identiche quando si studia la funzione sintetica delle articolazioni.
Il metabolismo dei condrociti e la loro risposta ai fattori regolatori dipendono in modo significativo anche dall'età del donatore, dal suo sviluppo scheletrico e dalle condizioni delle articolazioni da cui vengono prelevate le cellule. Nei condrociti umani, si osserva una significativa diminuzione della risposta proliferativa con l'età. La diminuzione maggiore si osserva nei donatori di età compresa tra 40 e 50 anni e oltre i 60 anni. Inoltre, la gravità della risposta proliferativa ai fattori di crescita (ad esempio, FGF e TGF-beta) diminuisce con l'invecchiamento. Oltre ai cambiamenti quantitativi nella proliferazione dei condrociti, si verificano anche cambiamenti qualitativi. Le cellule di donatori giovani (10-20 anni) rispondono meglio al fattore di crescita derivato dalle piastrine (PDGF) rispetto al TGF-beta, mentre il contrario si osserva nelle cellule di donatori adulti. Diversi meccanismi vengono utilizzati per spiegare i cambiamenti dipendenti dall'età nella funzione sintetica dei condrociti e la loro risposta ai fattori di crescita. Tra questi rientrano una diminuzione del numero e dell'affinità dei recettori cellulari di superficie, modifiche nella sintesi e nella bioattività dei fattori di crescita e delle citochine e modificazioni dei segnali post-recettoriali.
Anche le condizioni patologiche delle articolazioni alterano la morfologia e l'attività metabolica dei condrociti. Pertanto, J. Kouri et al. (1996) hanno identificato tre sottopopolazioni di condrociti nella cartilagine in caso di osteoartrosi. I condrociti provenienti dalla parte superficiale e superiore della cartilagine media formano cluster e sintetizzano una maggiore quantità di proteoglicani e collagene. Il TGF-β e il fattore di crescita insulino-simile (IGF) sono in grado di stimolare la sintesi di proteoglicani da parte dei condrociti e di neutralizzare parzialmente gli effetti di IL-1 e TNF-α. Gli espianti di cartilagine affetta da osteoartrosi e i condrociti isolati dalla cartilagine di un paziente con osteoartrosi sono più sensibili alla stimolazione del TGF-β rispetto ai condrociti di cartilagine sana. Queste differenze sono molto probabilmente associate a cambiamenti fenotipici nei condrociti degli strati superiori della cartilagine articolare.
L'isolamento dei singoli condrociti si ottiene mediante trattamento sequenziale della matrice extracellulare (ECM) con enzimi proteolitici. Dopo il rilascio dalla ECM, le cellule isolate sono ideali per studiare la sintesi de novo dei componenti della matrice. Alcuni autori utilizzano solo collagenasi clostridiale, mentre altri preincubano la cartilagine con tripsina, pronasi, DNasi e/o ialuronidasi. Il numero di cellule isolate dipende dagli enzimi utilizzati. Pertanto, se trattati con la sola collagenasi, si possono ottenere 1,4-10⁻¹ condrociti da 1 g di tessuto, mentre se trattati con pronasi, ialuronidasi e collagenasi, 4,3-10⁻¹ . Se trattati con collagenasi, aggrecano, proteine, IL-6 e IL-8 rimangono nella coltura cellulare in quantità significativamente maggiori rispetto al trattamento sequenziale con enzimi diversi. Esistono diverse spiegazioni per queste differenze tra le due colture cellulari:
- I recettori cellulari sono danneggiati o inibiti dagli enzimi, il TGF-β inibisce la sintesi di DNA e proteoglicani nei condrociti appena isolati (giorno 1), mentre la sintesi di DNA e proteoglicani nei condrociti coltivati in monostrato (7 giorni) è stimolata dal TGF-β. Tuttavia, è necessario un periodo adeguato per la riespressione di questi componenti di membrana prima dell'inizio dell'esperimento.
- Le proteasi esogene possono interrompere l'interazione cellula-matrice mediata dalle integrine. La famiglia delle integrine promuove l'adesione dei condrociti alle molecole della matrice extracellulare (ECM) (Shakibaei M. et al., 1997). Questa interruzione può influenzare l'espressione dei geni della matrice.
- I residui dei componenti della matrice extracellulare possono regolare la funzione sintetica dei condrociti. Le integrine sono in grado di riconoscere i prodotti di degradazione dell'ECM, svolgendo così un ruolo importante nella riparazione tissutale dopo l'azione degli enzimi proteolitici. T. Larsson et al. (1989) hanno riportato che l'aggiunta di proteoglicani intatti o frammentati a colture cellulari stimola la sintesi di proteine e proteoglicani. Tuttavia, un elevato livello di acido ialuronico causa una significativa riduzione dell'inclusione di solfati nella sintesi di proteoglicani da parte dei condrociti dell'embrione di pollo, dei condrociti maturi del maiale e delle cellule di condrosarcoma di ratto. Inoltre, l'acido ialuronico è un inibitore del rilascio di proteoglicani dalle cellule anche in presenza di IL-1b, TNF-α, FGF, il che indica una contrazione della prima attività biologica di fattori di crescita e citochine. L'esatto meccanismo d'azione dell'acido ialuronico rimane poco chiaro; È noto che i condrociti contengono un recettore per l'acido ialuronico associato ai filamenti di actina del citosol. Il legame dell'acido ialuronico al suo recettore stimola la fosforilazione proteica. Pertanto, questi dati dimostrano la modulazione della funzione metabolica dei condrociti da parte di molecole frammentate o native di proteine della matrice attraverso l'attivazione dei recettori di membrana cellulare.
- La rapida stimolazione della sintesi proteica della matrice da parte dei condrociti da parte degli enzimi può essere una conseguenza di cambiamenti nella forma dei condrociti e/o nella riorganizzazione del citoscheletro.
- Alcune citochine (ad esempio, IL-8) e fattori di crescita (ad esempio, IGF-1, TGF-β) sono sequestrati nella matrice extracellulare (ECM). L'esempio più noto è il legame del TGF-β alla decorina, che si traduce in una ridotta capacità del primo di indurre la crescita cellulare nelle cellule ovariche di criceto cinese. La scoperta che il contenuto di decorina nella cartilagine aumenta con l'età suggerisce una diminuzione della biodisponibilità del TGF-β con l'invecchiamento. Fattori di crescita e citochine possono essere rilasciati dai detriti della matrice durante la coltura e successivamente modulare la funzione dei condrociti.
Coltura monostrato di condrociti
Il fenotipo differenziato dei condrociti è caratterizzato principalmente dalla sintesi di collagene di tipo II e di proteoglicani tessuto-specifici, nonché da un basso livello di attività mitotica. È stato dimostrato che con la coltivazione cellulare prolungata in monostrato, così come dopo ripetuti passaggi cellulari, i condrociti perdono la loro forma sferica e acquisiscono una forma allungata, simile a quella dei fibroblasti. Con tale metaplasia fibroblastica, anche la funzione sintetica delle cellule viene modificata, caratterizzata da una progressiva diminuzione della sintesi di collageni di tipo II, IX e XI e da un aumento della sintesi di collageni di tipo I, III e Y. Piccoli proteoglicani non aggregati vengono sintetizzati grazie all'aggrecano funzionale. La sintesi di catepsina B ed L è estremamente bassa nelle cellule differenziate, ma aumenta durante il processo di perdita di differenziazione. La collagenasi-1 è espressa nei condrociti differenziati; con la coltivazione prolungata, la sua espressione diminuisce, mentre aumenta la produzione di inibitori tissutali delle metalloproteasi (TIMP).
I condrociti differenziati riesprimono il collagene del fenotipo differenziato quando trasferiti da un monostrato a una coltura sospesa. Il processo di differenziazione è probabilmente correlato alla forma cellulare. Questa proprietà viene regolarmente utilizzata dai ricercatori che studiano innesti difettosi con condrociti autologhi. Un piccolo numero di cellule ottenute da materiale bioptico può essere espanso in coltura monostrato e quindi reintrodotto in una matrice tridimensionale prima del trapianto. La riespressione di un fenotipo specifico da parte di condrociti dedifferenziati trasferiti in coltura di agarosio può essere stimolata da TGF-β, complesso osseina-idrossiapatite e acido ascorbico.
In risposta a fattori di crescita e citochine, i condrociti vengono modificati durante il processo di differenziazione. La risposta cellulare a citochine e fattori di crescita differisce tra condrociti indifferenziati e differenziati. L'IL-1 stimola la proliferazione dei fibroblasti, mentre la crescita dei condrociti indifferenziati è inibita dall'IL-1. La sintesi del DNA è stimolata dall'IGF-1 nei condrociti allungati ma non appiattiti. Nei condrociti differenziati, gli effetti stimolatori di IL-1β e TNF-α sulla produzione di procollagenasi sono più pronunciati rispetto ai condrociti indifferenziati.
Coltivazione di condrociti
La coltura di condrociti in sospensione in un mezzo liquido o in una matrice tridimensionale naturale o sintetica stabilizza il fenotipo condrocitario. Le cellule mantengono la loro forma sferica e sintetizzano proteine tessuto-specifiche. La coltura di condrociti in sospensione è generalmente raccomandata per studiare la formazione di una nuova matrice pericellulare. Le colture di condrociti in polimeri assorbenti sintetici o naturali vengono utilizzate per l'impianto di cellule in difetti cartilaginei al fine di stimolare la rigenerazione del tessuto cartilagineo articolare. Il mezzo sintetico o naturale per l'impianto di cellule deve soddisfare una serie di requisiti:
- gli impianti devono avere una struttura porosa per l'adesione e la crescita cellulare,
- né il polimero stesso né i suoi prodotti di degradazione dovrebbero causare infiammazioni o reazioni tossiche quando impiantati in vivo,
- il supporto dell'innesto deve avere la capacità di legarsi alla cartilagine adiacente o all'osso subcondrale,
- la matrice naturale o sintetica deve avere la capacità di assorbire, la sua degradazione deve essere bilanciata dalla rigenerazione dei tessuti,
- per facilitare la riparazione della cartilagine, la struttura chimica e l'architettura dei pori della matrice devono facilitare il mantenimento del fenotipo cellulare e la sintesi di proteine tessuto-specifiche da parte dei condrociti in essa collocati,
- Durante l'impianto in vivo è necessario studiare le proprietà meccaniche della matrice sintetica o naturale.
 [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
[ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
Sospensione di condrociti in fase liquida
L'adesione delle cellule ai vasi di plastica in cui vengono coltivati i condrociti può essere prevenuta rivestendone le pareti con una soluzione di metilcellulosa, agarosio, idrogel (poli-2-idrossietilmetacrilato) o una miscela di collagene e agarosio. In queste condizioni, i condrociti formano cluster e sintetizzano principalmente aggrecani e collageni tessuto-specifici (tipi II, IX, XI). Si riscontrano solitamente due tipi di cellule. Le cellule situate al centro mantengono una forma sferica e sono circondate da una matrice extracellulare ben sviluppata, come confermato da studi istochimici e ultrastrutturali. Alla periferia, i condrociti presentano contorni discoidi e sono circondati da una matrice extracellulare sparsa; si sa poco sulle caratteristiche funzionali di queste cellule.
È possibile coltivare condrociti su microcarrier mantenuti in sospensione; come microcarrier vengono utilizzate perle di destrano (cytodex), perle di destrano rivestite di collagene (cytodex III) e microsfere non porose di collagene di tipo I (cellagen). In queste condizioni di coltura, i condrociti si attaccano alla superficie del microcarrier, mantengono la loro forma sferica e producono un materiale simile alla matrice. Inoltre, l'uso di cellulagen promuove la proliferazione dei condrociti e la riespressione del fenotipo normale. Pertanto, la coltura di condrociti su microsfere di cellulagen può essere utilizzata per ripristinare il fenotipo cellulare prima del trapianto.
Un altro metodo per coltivare sospensioni di condrociti in un mezzo liquido è la loro coltura sotto forma di sfere dense costituite da cellule (0,5-1 * 10 b ), ottenute per centrifugazione. Tali condrociti sono in grado di produrre una matrice contenente un'elevata quantità di proteoglicani, collagene di tipo II, ma non di collagene di tipo I, come confermato da metodi istologici, immunoistochimici e quantitativi.
Sospensione di condrociti nella matrice extracellulare naturale
I condrociti possono essere coltivati in sospensione in una matrice tridimensionale (agar morbido, agarosio, gel o spugna di collagene, acido ialuronico, colla di fibrina, sfere di alginato).
I condrociti coltivati in agarosio mantengono il loro fenotipo normale e sintetizzano collagene di tipo II e aggregati di aggrecani tessuto-specifici. Quando coltivati in agarosio, i proteoglicani sintetizzati dalla cellula vengono rilasciati nel terreno di coltura per 50 giorni. A titolo di confronto, in una coltura monostrato, la fase cellulare è sovraccarica di glicosamminoglicani già nei primi 5-6 giorni di coltura; quando coltivati nel terreno di coltura, dopo un aumento della sintesi e del rilascio di glicosamminoglicani nei primi 8-10 giorni, si verifica una loro diminuzione tempo-dipendente. Tuttavia, il comportamento dei condrociti coltivati in agarosio differisce da quello in vivo. In agarosio, un gran numero di aggregati di aggrecani sintetizzati contiene molecole più piccole e meno molecole rispetto a quelle in vivo. Il TGF-β stimola la sintesi di proteoglicani nell'espianto, ma riduce la sintesi di aggrecani in agarosio.
L'alginato è un polisaccaride lineare ottenuto da alghe brune. In presenza di cationi bivalenti, come gli ioni Ca2 +, questo polimero si trasforma in un gel. Ogni condrocita intrappolato nell'alginato è circondato da una matrice di polisaccaridi a carica negativa, i cui pori sono paragonabili a quelli della cartilagine ialina. La matrice formata dai condrociti nelle microsfere di alginato è costituita da due compartimenti: un sottile strato di matrice associata alle cellule corrispondente alla matrice pericellulare e territoriale della cartilagine articolare e una matrice più distante equivalente a quella interterritoriale del tessuto nativo. Al trentesimo giorno di coltura, il volume relativo e assoluto occupato dalle cellule e da ciascuno dei due compartimenti in una microsferula di alginato è quasi completamente identico a quello della cartilagine nativa. Per quasi 30 giorni, i condrociti mantengono la loro forma sferica e producono aggrecano, le cui proprietà idrodinamiche sono simili a quelle delle molecole di aggrecano presenti nella matrice cartilaginea articolare, così come delle molecole di collagene di tipo II, IX e XI. Allo stesso tempo, come in altre colture in sospensione, sulla superficie delle perle di alginato sono presenti cellule appiattite, che producono una piccola quantità di molecole di collagene di tipo I, che vengono rilasciate direttamente nel terreno di coltura e non vengono incorporate nella matrice extracellulare (ECM). Nelle perle di alginato si osserva una moderata proliferazione di condrociti. Dopo 8 mesi di coltura in gel di alginato, i condrociti maturi non perdono attività metabolica e continuano a sintetizzare collagene di tipo II e aggrecano tessuto-specifici.
H. Tanaka et al. (1984) hanno studiato le proprietà di diffusione di varie molecole naturali in alginato e hanno scoperto che le molecole di peso superiore a 70 kDa non diffondevano attraverso l'alginato. Pertanto, la coltura cellulare in alginato è adatta allo studio della regolazione della biosintesi della matrice e dell'organizzazione della matrice extracellulare (ECM). La disponibilità di cellule coltivate in alginato consente di studiare l'azione dei fattori regolatori peptidici e degli agenti farmacologici a livello trascrizionale, post-trascrizionale e traduzionale.
I condrociti vengono anche coltivati in una matrice di fibre di collagene di tipo I e II. S. Nehrer et al. (1997) hanno confrontato il funzionamento dei condrociti canini in matrici porose di polimeri di collagene-proteoglicani contenenti collageni di tipo diverso. Hanno riscontrato importanti differenze nella morfologia della funzione biosintetica dei condrociti coltivati in matrici di collagene contenenti collagene di tipo I e II. Le cellule in una matrice di collagene di tipo II hanno mantenuto la loro forma sferica, mentre in quella di collagene di tipo I hanno mostrato una morfologia simile a quella dei fibroblasti. Inoltre, in una matrice di collagene di tipo II, i condrociti hanno prodotto una maggiore quantità di glicosaminoglicani. J. van Susante et al. (1995) hanno confrontato le proprietà dei condrociti coltivati in gel di alginato e di collagene (di tipo I). Gli autori hanno riscontrato un aumento significativo del numero di cellule nel gel di collagene, ma dal sesto giorno di coltivazione le cellule hanno perso il loro fenotipo caratteristico, trasformandosi in cellule simili ai fibroblasti. Nel gel di alginato è stata osservata una diminuzione del numero di cellule, ma i condrociti hanno mantenuto il loro fenotipo normale. Nel gel di collagene, il numero di proteoglicani per cellula era significativamente più alto rispetto all'alginato, ma nel gel è stata osservata una diminuzione nella sintesi di elementi della matrice, a partire dal sesto giorno di coltivazione, mentre nell'alginato la sintesi ha continuato ad aumentare.
La matrice di fibrina solida tridimensionale è una sostanza naturale che supporta i condrociti in essa sospesi in un fenotipo differenziato. La matrice di fibrina tridimensionale può anche essere utilizzata come vettore nel trapianto di condrociti. I vantaggi della fibrina sono l'assenza di citotossicità, la capacità di riempire gli spazi e la capacità adesiva. Studi istologici e biochimici, l'autoradiografia e la microscopia elettronica hanno dimostrato che i condrociti nel gel di fibrina mantengono la loro morfologia, proliferano e producono matrice anche dopo 2 settimane di coltivazione. Tuttavia, G. Homminga et al. (1993) hanno riportato che la disintegrazione della fibrina inizia dopo 3 giorni di coltivazione e la dedifferenziazione dei condrociti progredisce.
Sospensione di condrociti in ECM artificiale (sintetica)
Gli impianti cartilaginei per la chirurgia ricostruttiva o ortopedica possono essere ottenuti coltivando condrociti isolati in vitro in una matrice sintetica biocompatibile.
I condrociti coltivati in acido poliglicolico proliferano e mantengono morfologia e fenotipo normali per 8 settimane. Il complesso condrociti-acido poliglicolico è costituito da cellule, glicosamminoglicani, collageni e presenta una capsula di collagene esterna. Tuttavia, tali impianti contengono due tipi di molecole di collagene: I e II. Gli impianti ottenuti da condrociti dedifferenziati attraverso una serie di passaggi presentano una quantità maggiore di glicosamminoglicani e collageni rispetto agli impianti ottenuti da condrociti principalmente indifferenziati.
L. Freed et al. (1993b) hanno confrontato il comportamento di colture di condrociti umani e bovini in acido poliglicolico fibroso (FPGA) e acido polilattico poroso (PPLA). Dopo 6-8 settimane di coltura di condrociti bovini in FPGA o PPLA, gli autori hanno osservato la proliferazione cellulare e la rigenerazione della matrice cartilaginea. In FPGA, i condrociti avevano una forma sferica e si trovavano in lacune circondate da una matrice cartilaginea. Dopo 8 settimane di coltura in vitro, il tessuto rigenerato conteneva fino al 50% di sostanza secca (4% di massa cellulare, 15% di glicosaminoglicani e 31% di collageni). In PPLA, le cellule avevano una forma fusiforme e una piccola quantità di glicosaminoglicani e collagene. In FPGA, la crescita cellulare era 2 volte più intensa che in PPLA. In vivo, i condrociti coltivati in VPGK e PPLC hanno prodotto tessuto istologicamente simile alla cartilagine entro 1-6 mesi. Gli impianti contenevano glicosaminoglicani e collageni di tipo I e II.
Condrociti fetali bovini sono stati coltivati in polietilene poroso ad alta densità, idrofobico e idrofilo. Dopo 7 giorni di incubazione in entrambi i substrati, le cellule hanno mantenuto una forma sferica e contenevano principalmente collagene di tipo II. Dopo 21 giorni di coltivazione, si è scoperto che la matrice idrofila conteneva più collagene di tipo II rispetto a quella idrofobica.
Tessuto cartilagineo può essere ottenuto anche coltivando in monostrato su filtri Millicell-CM. Il pre-rivestimento dei filtri con collagene è necessario per l'adesione delle condroitine. L'esame istologico della coltura dimostra l'accumulo di condrociti nella matrice extracellulare (ECM) contenente proteoglicani e collagene di tipo II. Il collagene di tipo I non è stato rilevato in tale coltura. I condrociti nel tessuto cartilagineo ottenuto hanno una forma sferica, ma sulla superficie del tessuto sono leggermente appiattiti. Lo spessore del tessuto neoformato è aumentato nel tempo e dipendeva dalla densità iniziale del monostrato cellulare. In condizioni di coltura ottimali, lo spessore del tessuto cartilagineo ha raggiunto i 110 μm, e l'organizzazione delle sue cellule e del collagene in strati superficiali e profondi è simile a quella della cartilagine articolare. La matrice extracellulare (ECM) contiene circa 3 volte più collagene e proteoglicani. Dopo 2 settimane di coltura, è stato osservato un accumulo di matrice, che ha consentito l'estrazione del tessuto dal filtro e il suo utilizzo per il trapianto.
Sims et al. (1996) hanno studiato la coltura di condrociti in gel di ossido di polietilene, una matrice polimerica incapsulata che consente il trasferimento di un gran numero di cellule per iniezione. Sei settimane dopo l'iniezione nel tessuto sottocutaneo di topi atimici, si è formata nuova cartilagine, caratterizzata morfologicamente da un'opalescenza bianca simile alla cartilagine ialina. I dati istologici e biochimici hanno indicato la presenza di condrociti in proliferazione attiva, produttori di ECM.
Spiegazione
L'espianto di tessuto cartilagineo viene utilizzato per studiare i processi di ana- e catabolismo in esso contenuti, il mantenimento dell'omeostasi, il riassorbimento e la riparazione. I condrociti negli espianti cartilaginei mantengono un fenotipo normale e una composizione della matrice extracellulare (ECM) simile a quella della cartilagine articolare in vivo. Dopo 5 giorni di coltura in presenza di siero, si ottiene un livello costante di sintesi e processi di degradazione naturali. Il riassorbimento tissutale può essere accelerato nella coltura principale e in coltura con l'aggiunta di siero utilizzando diversi agenti, come IL-IB, TNF-α, lipopolisaccaridi batterici, derivati dell'acido retinoico o radicali dell'ossigeno attivo. Per studiare la riparazione della cartilagine, il danno viene indotto da mediatori infiammatori solubili (H₂O₂ , IL 1, TNF- α ) o dalla rottura fisica della matrice.
Il metodo di coltura organotipica è un modello per studi in vitro sugli effetti di fattori esterni isolati sui condrociti e sulla matrice extracellulare circostante. In vivo, i condrociti sono localizzati in modo sparso nella matrice extracellulare (ECM) e non sono in contatto tra loro. La coltura di espianti di cartilagine articolare preserva questa organizzazione strutturale, nonché le interazioni specifiche tra i condrociti e l'ambiente extracellulare circostante. Questo modello viene utilizzato anche per studiare gli effetti di stress meccanico, agenti farmacologici, fattori di crescita, citochine e ormoni sul metabolismo cartilagineo.
Un altro vantaggio dell'espianto di tessuto cartilagineo è l'assenza di danni ai condrociti sotto l'azione di enzimi proteolitici o fattori meccanici, inevitabile durante l'isolamento cellulare. I recettori e altre proteine di membrana e glicoproteine sono protetti da fattori dannosi.
[ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ]
Coltura di Chondron
Il condrone è un'unità strutturale, funzionale e metabolica della cartilagine articolare costituita da un condrocita, dalla sua matrice pericellulare e dalla sua capsula filamentosa compatta, responsabile dell'omeostasi della matrice. I condroni vengono estratti meccanicamente dalla cartilagine e raccolti mediante diverse omogeneizzazioni successive a bassa velocità. I condroni isolati da zone di diversa profondità cartilaginea possono essere suddivisi in quattro categorie: condrone singolo, condrone appaiato, condrone multiplo (tre o più) disposto linearmente (colonne di condroni) e cluster di condroni.
I condri singoli si trovano solitamente negli strati intermedi della cartilagine intatta, i condri appaiati si trovano al confine tra gli strati intermedi e profondi, mentre i condri multipli disposti linearmente sono tipici degli strati profondi della cartilagine intatta. Infine, i cluster di condri sono costituiti da gruppi di condri singoli e appaiati organizzati casualmente, che mantengono uno stato aggregato dopo l'omogeneizzazione. I cluster di condri sono grandi frammenti di cartilagine, solitamente contenenti diversi condri e fibrille di collagene disposte radialmente, ovvero una tipica organizzazione caratteristica degli strati profondi della matrice. I condri sono immobilizzati in agarosio trasparente, il che consente di studiarne la struttura, la composizione molecolare e l'attività metabolica. Il sistema condrociti-agarosio è considerato un micromodello di cartilagine, che differisce dal tradizionale sistema condrociti-agarosio in quanto il microambiente naturale viene preservato e non è necessario sintetizzarlo e assemblarlo. La coltura di condroni è un modello per studiare le interazioni tra cellule e matrice nella cartilagine articolare in condizioni normali e patologiche.
[ 22 ], [ 23 ], [ 24 ], [ 25 ], [ 26 ], [ 27 ]
Coltura di condrociti immortali
Virus contenenti DNA ricombinante o oncogeni in grado di rendere una cellula "immortale" vengono utilizzati per creare linee cellulari permanenti. I condrociti immortali hanno la capacità di proliferare all'infinito mantenendo un fenotipo stabile. F. Mallein-Gerin et al. (1995) hanno dimostrato che l'oncogene SV40T induce la proliferazione dei condrociti murini, che continuano a esprimere stabilmente i tipi di collagene II, IX e XI, nonché l'aggrecano articolare e la proteina legante. Tuttavia, una tale linea cellulare acquisisce la capacità di sintetizzare il collagene di tipo I quando viene coltivata in una coltura monostrato o in un gel di agarosio.
W. Horton et al. (1988) hanno descritto una linea di cellule immortali con un basso livello di espressione dell'mRNA del collagene di tipo II. Queste cellule sono state ottenute trasformandole con un retrovirus murino contenente gli oncogeni I-myc e y-ra. Questo tipo di cellule rappresenta un modello unico per lo studio delle interazioni della matrice articolare in assenza di collagene di tipo II, nonché della regolazione della sintesi di collagene di tipo II.
La coltura di condropriti con geni mutati o deleti è un modello utile per studiarne la funzione fisiologica. Questo modello è particolarmente adatto per studiare il ruolo di specifiche molecole nell'organizzazione della matrice cartilaginea o per indagare gli effetti di vari fattori regolatori sul metabolismo cartilagineo. I condrociti con un gene deleto per il collagene di tipo IX sintetizzano fibrille di collagene più larghe del normale, a indicare che il collagene di tipo IX regola il diametro delle fibrille. Come osservato nel Capitolo 1, è stata recentemente scoperta una mutazione nel gene COLAI, che codifica per il collagene di tipo II, in famiglie con osteoartrite generalizzata primaria. Per studiare l'effetto del collagene di tipo II mutante sulla matrice articolare, R. Dharmrvaram et al. (1997) hanno trasfettato (infettato con acido nucleico estraneo) COL 2 AI difettoso (l'arginina in posizione 519 è sostituita da cisteina) in condrociti fetali umani in vitro.
Sistema di co-coltura. Nell'articolazione, la cartilagine interagisce con altri tipi di cellule contenute nella membrana sinoviale, nel liquido sinoviale, nei legamenti e nell'osso subcondrale. Il metabolismo dei condrociti può essere influenzato da vari fattori solubili sintetizzati dalle cellule elencate. Pertanto, nell'artrite, la cartilagine articolare viene distrutta dagli enzimi proteolitici e dai radicali liberi prodotti dalle cellule sinoviali. Pertanto, sono stati sviluppati modelli per studiare le complesse interazioni tra cartilagine e tessuti circostanti, chiamati co-colture.
S. Lacombe-Gleise et al. (1995) hanno coltivato condrociti e osteoblasti di coniglio in un sistema di cocoltura (COSTAR) in cui le cellule erano separate da una membrana microporosa (0,4 μm) che consentiva lo scambio tra i due tipi cellulari senza alcun contatto diretto. Questo studio ha dimostrato la capacità degli osteoblasti di stimolare la crescita dei condrociti tramite mediatori solubili.
AM Malfait e coautori (1994) hanno studiato la relazione tra monociti e condrociti del sangue periferico. Questo modello è utile per studiare i processi mediati dalle citochine nelle artropatie infiammatorie (artrite reumatoide, spondiloartriti sieronegative, ecc.). Gli autori del modello hanno separato le cellule con una membrana legante le proteine con pori di 0,4 μm di diametro. Lo studio ha dimostrato che i monociti stimolati con lipopolisaccaride producevano IL-1 e TNF-α, che inibivano la sintesi di aggrecano da parte dei condrociti e contribuivano alla degradazione degli aggregati di aggrecano già sintetizzati.
K. Tada et al. (1994) hanno creato un modello di cocoltura in cui cellule endoteliali in un gel di collagene (tipo I) venivano collocate in una camera interna separata dalla camera esterna, con condrociti inseriti al suo interno tramite un filtro con pori di 0,4 μm. In uno stato di completo isolamento dalla camera esterna, le cellule endoteliali umane formavano tubuli in un gel di collagene in presenza di EGF o TGF-α. Quando entrambi i tipi di cellule venivano coltivati simultaneamente, la formazione di tubuli TGF-α-dipendente da parte delle cellule endoteliali veniva inibita. L'inibizione di questo processo da parte dei condrociti veniva parzialmente eliminata dagli anticorpi anti-TGF-β. Si può supporre che il TGF-β prodotto dai condrociti inibisca la vascolarizzazione della cartilagine stessa.
S. Groot et al. (1994) hanno coltivato simultaneamente condrociti provenienti dalle zone ipertrofiche e proliferative dell'osso di un feto di topo di 16 giorni con frammenti di tessuto cerebrale. Dopo 4 giorni di coltura, si è osservata la transdifferenziazione dei condrociti in osteoblasti e l'inizio della formazione di osteoide. Dopo 11 giorni di coltura, parte della cartilagine è stata sostituita da tessuto osseo e la matrice ossea è risultata parzialmente calcificata. Alcuni neuropeptidi e neurotrasmettitori prodotti dal tessuto cerebrale influenzano il metabolismo degli osteoblasti o possiedono recettori per essi. Tra questi figurano la noradrenalina, il peptide intestinale vasoattivo, il peptide correlato al gene della calcitonina, la sostanza P e la somatostatina. Frammenti di tessuto cerebrale coltivati insieme ai condrociti possono produrre alcuni dei fattori elencati in grado di indurre il processo di transdifferenziazione dei condrociti in osteoblasti.
[ 28 ], [ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ]
L'influenza dei fattori esterni sulla coltura dei condrociti
L'influenza della tensione di ossigeno sul metabolismo dei condrociti
Nella maggior parte dei casi, le colture di condrociti si sviluppano in condizioni di tensione atmosferica dell'ossigeno. Tuttavia, è noto che in vivo i condrociti esistono in condizioni di ipossia e la tensione dell'ossigeno varia in diverse condizioni patologiche. Durante il processo di maturazione, si osservano cambiamenti significativi nell'afflusso di sangue alle epifisi. Poiché la vascolarizzazione varia nelle diverse zone della cartilagine di accrescimento, anche la tensione dell'ossigeno in esse varia. C. Brighton e R. Heppenstall (1971) hanno dimostrato che nella cartilagine tibiale dei conigli, la tensione dell'ossigeno nella zona ipertrofica è inferiore rispetto alla cartilagine circostante. Le misurazioni di alcuni parametri metabolici hanno mostrato che i condrociti sono in grado di rispondere rapidamente alle variazioni locali della concentrazione di ossigeno. Innanzitutto, a bassa tensione dell'ossigeno, il suo consumo da parte dei condrociti diminuisce. Con una diminuzione della tensione dell'ossigeno dal 21% allo 0,04%, aumenta l'utilizzo del glucosio, l'attività degli enzimi glicolitici e la sintesi di acido lattico. Anche a bassa tensione di ossigeno, la quantità assoluta di ATP, ADP e AMP rimane stabile. Questi dati indicano che il metabolismo dei condrociti è finalizzato alla massima conservazione dell'energia. Tuttavia, l'attività di sintesi, e quindi i processi di riparazione, variano in condizioni di ipossia.
Un'elevata tensione di ossigeno influisce anche sul metabolismo dei condrociti, causando una riduzione della sintesi di proteoglicani e DNA e la degradazione della matrice cartilaginea. Questi effetti sono solitamente accompagnati dalla produzione di radicali liberi dell'ossigeno.
L'influenza della concentrazione ionica e della pressione osmotica dell'ambiente sulla funzione dei condrociti
Nella cartilagine nativa, la concentrazione di ioni differisce significativamente da quella di altri tessuti: il contenuto di sodio nel mezzo extracellulare è di 250-350 mmol e la sua osmolarità è di 350-450 mosmol. Quando i condrociti vengono isolati dalla matrice extracellulare (ECM) e incubati in terreni standard (DMEM - Dulbecco's Minimal Essential Medium), con osmolarità di 250-280,7 mosmol), l'ambiente circostante le cellule cambia drasticamente. Inoltre, la concentrazione di calcio e potassio nei terreni standard è significativamente inferiore rispetto al tessuto nativo e la concentrazione di anioni è significativamente maggiore.
L'aggiunta di saccarosio al terreno di coltura ne aumenta l'osmolarità e induce un aumento intracellulare transitorio della concentrazione di anioni H + e calcio nel citosol. Tali cambiamenti intracellulari possono influenzare i processi di differenziazione dei condrociti e la loro attività metabolica. J. Urban et al. (1993) hanno scoperto che l'incorporazione di 35 8-solfato e 3 H-prolina da parte di condrociti isolati incubati in DMEM standard per 2-4 ore era solo il 10% di quella presente nel tessuto nativo. L'intensità di sintesi raggiungeva il massimo a un'osmolarità del terreno di coltura extracellulare di 350-400 mosmol sia nei condrociti appena isolati che negli espianti di tessuto cartilagineo. Inoltre, il volume dei condrociti aumentava del 30-40% dopo aver posto le cellule isolate in DMEM standard con l'osmolarità specificata. Tuttavia, quando i condrociti vengono coltivati in condizioni di osmolarità non fisiologica per 12-16 ore, le cellule si adattano alle nuove condizioni, riducendo l'intensità della biosintesi in proporzione allo spostamento dell'osmolarità dell'ambiente extracellulare.
P. Borgetti et al. (1995) hanno studiato l'effetto dell'osmolarità del mezzo extracellulare sulla crescita, la morfologia e la biosintesi dei condrociti di maiale. Gli autori hanno dimostrato caratteristiche biochimiche e morfologiche simili nei condrociti coltivati in terreni con osmolarità di 0,28 e 0,38 mosmol. A un'osmolarità media di 0,48 mosmol, è stata osservata una diminuzione della proliferazione cellulare e della sintesi proteica durante le prime 4-6 ore di coltura, ma questi parametri si sono successivamente ripristinati, raggiungendo infine i valori di controllo. Quando i condrociti venivano coltivati in un terreno con un'osmolarità di 0,58 mosmol, le cellule perdevano la capacità di mantenere l'intensità fisiologica dei processi proliferativi e, dopo 6 giorni, il numero di condrociti risultava significativamente ridotto. A un'osmolarità media di 0,58 mosmol, è stata osservata una profonda inibizione della sintesi proteica. Inoltre, quando coltivati in terreni con un'osmolarità di 0,28-0,38 mOsm, i condrociti mantengono il loro fenotipo fisiologico; a osmolarità più elevate (0,48-0,58 mOsm), si verificano cambiamenti significativi nella morfologia cellulare, che si manifestano con la perdita del fenotipo caratteristico, la trasformazione dei condrociti in cellule fibroblasto-simili e la perdita della capacità delle cellule di assemblare proteoglicani della matrice. I risultati di questo studio indicano la capacità dei condrociti di rispondere a fluttuazioni limitate dell'osmolarità dell'ambiente extracellulare.
Anche le variazioni nella concentrazione di altri ioni possono influenzare i processi di biosintesi nei condrociti. Pertanto, il grado di incorporazione di 35 S (solfato) aumenta della metà con un aumento della concentrazione di ioni potassio da 5 mmol (la concentrazione nel mezzo DM EM standard) a 10 mmol (la concentrazione nell'ECM in vivo). Concentrazioni di calcio inferiori a 0,5 mmol hanno promosso la produzione di collagene da parte dei condrociti bovini maturi, mentre una concentrazione di 1-2 mmol (corrispondente alla concentrazione nel mezzo DM EM standard) ha causato una significativa diminuzione della sintesi di collagene. Un moderato aumento della biosintesi è stato osservato ad alti livelli di calcio (2-10 mmol). Vari cationi partecipano all'adesione dei condrociti alle proteine dell'ECM. Pertanto, gli ioni magnesio e manganese forniscono l'adesione alla fibronectina e al collagene di tipo II, mentre gli ioni calcio non partecipano all'adesione dei condrociti alle proteine. Pertanto, i risultati degli studi descritti indicano l'influenza delle variazioni degli ioni extracellulari di potassio, sodio, calcio e dell'osmolarità del mezzo sulla funzione biosintetica dei condrociti incubati in mezzi standard.
Effetto dello stress meccanico sul metabolismo dei condrociti
L'immobilizzazione articolare causa un'atrofia reversibile della cartilagine, che indica la necessità di stimoli meccanici per i normali processi metabolici nella matrice extracellulare (ECM). Nella maggior parte dei casi, i modelli di coltura cellulare utilizzati sono in condizioni di pressione atmosferica normale. M. Wright et al. (1996) hanno dimostrato che l'ambiente meccanico influenza il metabolismo dei condrociti; la risposta cellulare dipende dall'intensità e dalla frequenza del carico compressivo. Esperimenti con carico su espianti di cartilagine articolare intatta in vitro hanno dimostrato una diminuzione della sintesi di proteine e proteoglicani sotto l'azione del carico statico, mentre il carico dinamico stimola questi processi. I meccanismi esatti dell'effetto del carico meccanico sulla cartilagine sono complessi e sono probabilmente associati alla deformazione cellulare, alla pressione idrostatica, alla pressione osmotica, al potenziale elettrico e ai recettori cellulari di superficie per le molecole della matrice. Per studiare l'effetto di ciascuno di questi parametri, è necessario creare un sistema in cui ciascun parametro possa essere variato in modo indipendente. Ad esempio, la coltura di espianto non è adatta allo studio della deformazione cellulare, ma può essere utilizzata per studiare l'effetto generale della pressione sull'attività metabolica dei condrociti. La compressione della cartilagine porta alla deformazione cellulare ed è anche accompagnata dalla comparsa di un gradiente di pressione idrostatica, potenziale elettrico, flusso di fluidi e variazioni di parametri fisico-chimici come il contenuto d'acqua nella matrice, la densità di carica elettrica e il livello di pressione osmotica. La deformazione cellulare può essere studiata utilizzando condrociti isolati immersi in gel di agarosio o collagene.
Sono stati sviluppati diversi sistemi per studiare l'effetto della stimolazione meccanica sulla coltura di condrociti. Alcuni ricercatori utilizzano sistemi in cui la pressione viene applicata alla coltura cellulare attraverso la fase gassosa. Pertanto, JP Veldhuijzen et al. (1979), utilizzando una pressione di 13 kPa superiore a quella atmosferica a bassa frequenza (0,3 Hz) per 15 minuti, hanno osservato un aumento della sintesi di cAMP e proteoglicani e una diminuzione della sintesi di DNA. R. Smith et al. (1996) hanno dimostrato che l'esposizione intermittente di una coltura di condrociti bovini primari a pressione idrostatica (10 MPa) a una frequenza di 1 Hz per 4 ore ha causato un aumento della sintesi di aggrecano e collagene di tipo II, mentre la pressione costante non ha influenzato questi processi. Utilizzando un sistema simile, Wright et al. (1996) hanno riportato che la pressione ciclica sulla coltura cellulare è associata all'iperpolarizzazione della membrana cellulare dei condrociti e all'attivazione dei canali del potassio Ca2 + -dipendenti. Pertanto, gli effetti della pressione ciclica sono mediati dai canali ionici attivati dallo stiramento nella membrana dei condrociti. La risposta dei condrociti alla pressione idrostatica dipende dalle condizioni di coltura cellulare e dalla frequenza del carico applicato. Pertanto, la pressione idrostatica ciclica (5 MPa) riduce l'incorporazione di solfato nel monostrato di condrociti a frequenze di 0,05, 0,25 e 0,5 Hz, mentre a frequenze superiori a 0,5 Hz, l'incorporazione di solfato nell'espianto cartilagineo aumenta.
M. Bushmann et al. (1992) hanno riportato che i condrociti in gel di agarosio alterano la biosintesi in risposta a carichi meccanici statici e dinamici, allo stesso modo dell'organo intatto coltivato. Gli autori hanno scoperto che il carico meccanico genera uno stimolo iperosmotico con conseguente diminuzione del pH nei condrociti.
L'effetto dello stiramento meccanico può essere studiato su una coltura cellulare immersa in un gel. La forza di stiramento può essere creata utilizzando un vuoto controllato da computer. Quando il sistema è sotto un certo grado di vuoto, il fondo della capsula di Petri contenente la coltura cellulare viene esteso di una quantità nota; la deformazione è massima ai bordi del fondo della capsula e minima al centro. Lo stiramento viene trasmesso anche ai condrociti coltivati nella capsula di Petri. Utilizzando questo metodo, K. Holmvall et al. (1995) hanno dimostrato che nelle cellule di condrosarcoma coltivate in un gel di collagene (tipo II), l'espressione dell'mRNA di un'integrina β-β è aumentata. L' integrinaβ-β è in grado di legarsi al collagene di tipo II. È considerata un meccanorecettore, poiché interagisce con le proteine che legano l'actina, collegando così la matrice extracellulare (ECM) al citoscheletro.
L'effetto del pH sul metabolismo dei condrociti
Il pH del fluido interstiziale della matrice extracellulare (ECM) del tessuto cartilagineo è più acido rispetto ad altri tessuti. A. Maroudas (1980) ha determinato il pH della matrice cartilaginea articolare a 6,9. B. Diamant et al. (1966) hanno riscontrato un pH di 5,5 in condizioni patologiche. È noto che i condrociti vivono a bassa PO2, il che indica l'importante ruolo della glicolisi (95% di tutto il metabolismo del glucosio) nel metabolismo di queste cellule; la glicolisi è accompagnata dalla produzione di una grande quantità di acido lattico.
Oltre all'acidificazione dell'ambiente da parte dei prodotti della glicolisi, i componenti stessi della matrice sono di grande importanza. L'elevata quantità di carica negativa fissa sui proteoglicani modifica la composizione ionica extracellulare: si osserva un'alta concentrazione di cationi liberi (ad esempio, H +, Na +, K + ) e una bassa concentrazione di anioni (ad esempio, O₂, HCO₂). Inoltre, sotto l'influenza del carico meccanico, l'acqua viene espulsa dalla ECM, il che porta a un aumento della concentrazione di cariche negative fisse e all'attrazione di più cationi nella matrice. Ciò è accompagnato da una diminuzione del pH dell'ambiente extracellulare, che influenza il pH intracellulare, modificando così il metabolismo dei condrociti. R. Wilkin e A. Hall (1995) hanno studiato l'effetto del pH dell'ambiente extracellulare e intracellulare sulla biosintesi della matrice da parte di condrociti bovini isolati. Hanno osservato una duplice modificazione della sintesi della matrice con una diminuzione del pH. Una leggera diminuzione del pH (7,4
[ 34 ], [ 35 ], [ 36 ], [ 37 ], [ 38 ], [ 39 ], [ 40 ]
L'influenza della composizione del terreno di coltura sul metabolismo dei condrociti
Il terreno di coltura per i condrociti deve essere conforme alle condizioni sperimentali. Negli ultimi anni, il siero di vitello è stato utilizzato per ottimizzare le condizioni di coltura. Tuttavia, quando si utilizza il siero, è necessario tenere conto di alcuni punti importanti:
- crescita verso l'esterno di cellule dalla periferia del tessuto nelle colture di organi,
- variabilità nella composizione dei sieri di diverse serie,
- la presenza di componenti sconosciuti in essi,
- aumento del rischio di interferenze e artefatti quando si studia l'influenza di vari fattori biologici sull'attività metabolica delle cellule.
Un esempio di quest'ultimo è uno studio sull'effetto dell'EGF sui condrociti cartilaginei nei ratti. L'EGF ha stimolato l'incorporazione di 3 -H-timidina e un aumento del contenuto di DNA nella coltura. Questo effetto era più pronunciato a basse concentrazioni sieriche (<1%), ma a concentrazioni elevate (>7,5%) l'effetto scompariva.
È noto che i livelli di sintesi e degradazione nel DMEM integrato con siero di vitello sono significativamente aumentati rispetto alle condizioni in vivo. Le differenze tra il metabolismo in vivo e in vitro possono essere dovute a differenze tra il liquido sinoviale e il terreno di coltura delle cellule. Lee et al. (1997) hanno coltivato condrociti bovini giovani in agarosio utilizzando un terreno nutritivo contenente DMEM integrato con il 20% di siero di vitello e una grande quantità di liquido sinoviale allogenico normale. La presenza di liquido sinoviale nel terreno ha indotto un aumento della quantità di proteoglicani, fino all'80% della quantità totale di liquido sinoviale. Questi risultati indicano che il liquido sinoviale in coltura induce un livello di metabolismo simile a quello in vivo, con un elevato livello di sintesi di glicosaminoglicani e un basso livello di divisione cellulare.
G. Verbruggen et al. (1995) hanno dimostrato che la sintesi di 35 S-arrpeKaHa da parte di condrociti umani coltivati in agarosio in DMEM privo di siero era pari al 20-30% del livello di sintesi osservato in DMEM arricchito con siero di vitello al 10%. Gli autori hanno determinato in che misura IGF-1, IGF-2, TGF-R o insulina ripristinassero la produzione di aggrecani in terreno privo di siero. Gli autori hanno concluso che 100 ng/ml di insulina, IGF-1 o IGF-2 ripristinavano parzialmente la sintesi di aggrecani al 39-53% del livello di controllo. Non è stato osservato alcun sinergismo o accumulo con una combinazione dei fattori elencati. Allo stesso tempo, 10 ng/ml di TGF-R in presenza di 100 ng/ml di insulina stimolavano la sintesi di aggrecani al 90% o più del livello di riferimento. Infine, la transferrina sierica umana, da sola o in combinazione con insulina, non ha influenzato la sintesi di aggrecani. Quando il siero di vitello è stato sostituito con albumina sierica bovina, il contenuto di aggregati di aggrecani è diminuito significativamente. L'arricchimento del terreno di coltura con insulina, IGF o TGF-R ha parzialmente ripristinato la capacità delle cellule di produrre aggregati di aggrecani. Inoltre, IGF-1 e insulina sono in grado di mantenere l'omeostasi nelle colture cellulari. Dopo 40 giorni di coltivazione in un terreno arricchito con 10-20 ng/ml di IGF-1, la sintesi di proteoglicani si è mantenuta allo stesso livello o addirittura superiore rispetto al terreno contenente il 20% di siero di vitello. I processi catabolici sono proceduti più lentamente nel terreno arricchito con IGF-1 rispetto al terreno arricchito con una soluzione di albumina allo 0,1%, ma leggermente più velocemente nel terreno arricchito con il 20% di siero. Nelle colture a lunga vita, 20 ng/ml di IGF-1 mantengono stabile lo stato cellulare.
D. Lee et al. (1993) hanno confrontato l'effetto della composizione del terreno di coltura (DMEM, DMEM+20% siero di vitello, DMEM+20 ng/ml IGF-1) sulla sintesi del DNA in una coltura di espianto di tessuto cartilagineo, in una coltura monostrato e in sospensione di agarosio. Nella coltura in agarosio in presenza di siero, gli autori hanno osservato una tendenza dei condrociti a raggrupparsi in grandi cluster. Le cellule coltivate senza siero o con IGF-1 hanno mantenuto una forma rotonda in agarosio, si sono raccolte in piccoli gruppi, ma non hanno formato grandi aggregati. In un monostrato, la sintesi del DNA è stata significativamente maggiore nel terreno contenente siero rispetto al terreno arricchito con IGF-1; la sintesi del DNA in quest'ultimo è stata significativamente maggiore rispetto al terreno non arricchito. Non sono state riscontrate differenze nella sintesi del DNA quando i condrociti sono stati coltivati in sospensione di agarosio in un terreno non arricchito e in un terreno con IGF-1. Allo stesso tempo, la coltura di sospensioni di condrociti in agarosio in un terreno arricchito di siero è stata accompagnata da una maggiore incorporazione del radionucleotide 3 H-timidina rispetto ad altri terreni.
La vitamina C è necessaria per l'attivazione degli enzimi coinvolti nella formazione di una struttura elicoidale stabile delle fibrille di collagene. I condrociti carenti di acido ascorbico sintetizzano precursori del collagene non elicoidali sotto-idrossilati, che vengono secreti lentamente. La somministrazione di acido ascorbico (50 μg/ml) provoca l'idrossilazione dei collageni di tipo II e IX e la loro secrezione in quantità normali. L'aggiunta di vitamina C non ha influenzato il livello di sintesi dei proteoglicani. Pertanto, la secrezione di collagene è regolata indipendentemente dalla secrezione di proteoglicani.