Esperto medico dell'articolo

Nuove pubblicazioni
ipofisi
Ultima recensione: 04.07.2025

Tutti i contenuti di iLive sono revisionati o verificati da un punto di vista medico per garantire la massima precisione possibile.
Abbiamo linee guida rigorose in materia di sourcing e colleghiamo solo a siti di media affidabili, istituti di ricerca accademici e, ove possibile, studi rivisti dal punto di vista medico. Nota che i numeri tra parentesi ([1], [2], ecc.) Sono link cliccabili per questi studi.
Se ritieni che uno qualsiasi dei nostri contenuti sia impreciso, scaduto o comunque discutibile, selezionalo e premi Ctrl + Invio.
L'ipofisi (ipofisi, S. glandula pituitaria) è situata nella fossa ipofisaria della sella turcica dell'osso sfenoide ed è separata dalla cavità cranica da un processo della dura madre, formando il diaframma sellare. Attraverso l'apertura di questo diaframma, l'ipofisi è collegata all'infundibolo dell'ipotalamo del diencefalo. Le dimensioni trasversali dell'ipofisi sono 10-17 mm, quelle anteroposteriori 5-15 mm, quelle verticali 5-10 mm. La massa dell'ipofisi nell'uomo è di circa 0,5 g, nelle donne di 0,6 g. L'ipofisi è rivestita esternamente da una capsula.
In base allo sviluppo dell'ipofisi da due diversi rudimenti, nell'organo si distinguono due lobi: anteriore e posteriore. L'adenoipofisi, o lobo anteriore (adenohypophysis, s.lobus anterior), è più grande e costituisce il 70-80% della massa totale dell'ipofisi. È più densa del lobo posteriore. Nel lobo anteriore si distingue una parte distale (pars distalis), che occupa la parte anteriore della fossa ipofisaria, una parte intermedia (pars intermedia), situata al confine con il lobo posteriore, e una parte tuberosa (pars tuberalis), che si estende verso l'alto e si collega all'infundibolo dell'ipotalamo. A causa dell'abbondanza di vasi sanguigni, il lobo anteriore ha un colore giallo pallido con una sfumatura rossastra. Il parenchima dell'ipofisi anteriore è rappresentato da diversi tipi di cellule ghiandolari, tra i cui filamenti si trovano capillari sanguigni sinusoidali. Metà (50%) delle cellule dell'adenoipofisi sono adenociti cromofili, che presentano granuli a grana fine nel citoplasma che si colorano bene con i sali di cromo. Questi sono adenociti acidofili (40% di tutte le cellule dell'adenoipofisi) e adenociti basofili (10%). Gli adenociti basofili includono endocrinociti gonadotropi, corticotropi e tireotropi. Gli adenociti cromofobi sono piccoli, hanno un nucleo grande e una piccola quantità di citoplasma. Queste cellule sono considerate precursori degli adenociti cromofili. L'altro 50% delle cellule dell'adenoipofisi è costituito da adenociti cromofobi.
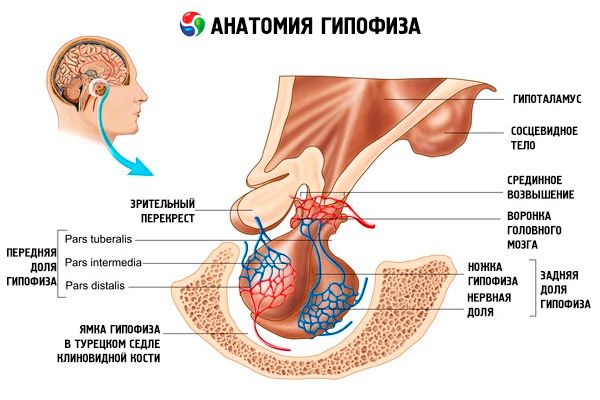
La neuroipofisi, o lobo posteriore (neuroipofisi, s.lobus posterior), è costituita dal lobo nervoso (lobus nervososus), situato nella parte posteriore della fossa ipofisaria, e dall'imbuto (infundibolo), situato dietro la parte tuberosa dell'adenoipofisi. Il lobo posteriore dell'ipofisi è formato da cellule neurogliali (cellule ipofisarie), fibre nervose che si diramano dai nuclei neurosecretori dell'ipotalamo alla neuroipofisi, e da corpuscoli neurosecretori.
L'ipofisi, tramite fibre nervose (vie) e vasi sanguigni, è funzionalmente connessa all'ipotalamo del diencefalo, che ne regola l'attività. L'ipofisi e l'ipotalamo, insieme alle loro connessioni neuroendocrine, vascolari e nervose, sono generalmente considerati il sistema ipotalamo-ipofisario.
Gli ormoni dell'ipofisi anteriore e posteriore influenzano molte funzioni dell'organismo, principalmente attraverso altre ghiandole endocrine. Nell'ipofisi anteriore, gli adenociti acidofili (cellule alfa) producono l'ormone somatotropo (ormone della crescita), che partecipa alla regolazione dei processi di crescita e sviluppo negli organismi giovani. Gli endocrinociti corticotropi secernono l'ormone adrenocorticotropo (ACTH), che stimola la secrezione di ormoni steroidei da parte delle ghiandole surrenali. Gli endocrinociti tireotropi secernono l'ormone tireotropo (TSH), che influenza lo sviluppo della tiroide e attiva la produzione dei suoi ormoni. Gli ormoni gonadotropi: follicolo-stimolante (FSH), luteinizzante (LH) e prolattina - influenzano la maturazione sessuale dell'organismo, regolano e stimolano lo sviluppo dei follicoli ovarici, l'ovulazione, la crescita delle ghiandole mammarie e la produzione di latte nelle donne, e il processo di spermatogenesi negli uomini. Questi ormoni sono prodotti dagli adenociti basofili (cellule beta). Qui vengono secreti anche i fattori lipotropici dell'ipofisi, che influenzano la mobilizzazione e l'utilizzo dei grassi nell'organismo. Nella parte intermedia del lobo anteriore viene prodotto l'ormone melanocita-stimolante, che controlla la formazione di pigmenti - melanina - nell'organismo.
Le cellule neurosecretorie dei nuclei sopraottico e paraventricolare nell'ipotalamo producono vasopressina e ossitocina. Questi ormoni vengono trasportati alle cellule dell'ipofisi posteriore lungo gli assoni che costituiscono il tratto ipotalamo-ipofisario. Dall'ipofisi posteriore, queste sostanze entrano nel sangue. L'ormone vasopressina ha un effetto vasocostrittore e antidiuretico, per il quale è anche chiamato ormone antidiuretico (ADH). L'ossitocina ha un effetto stimolante sulla contrattilità della muscolatura uterina, aumenta la secrezione di latte da parte della ghiandola mammaria in allattamento, inibisce lo sviluppo e la funzionalità del corpo luteo e influenza le alterazioni del tono della muscolatura liscia (non striata) del tratto gastrointestinale.
Sviluppo della ghiandola pituitaria
Il lobo anteriore dell'ipofisi si sviluppa dall'epitelio della parete dorsale della gola sotto forma di un'escrescenza anulare (tasca di Rathke). Questa protrusione ectodermica si sviluppa verso il fondo del futuro terzo ventricolo. Verso di essa, dalla superficie inferiore della seconda vescicola cerebrale (il futuro fondo del terzo ventricolo), si sviluppa un processo da cui si sviluppano il tubercolo grigio dell'infundibolo e il lobo posteriore dell'ipofisi.
Vasi e nervi della ghiandola pituitaria
Dalle arterie carotidi interne e dai vasi del circolo arterioso del cervello, le arterie ipofisarie superiore e inferiore si dirigono verso l'ipofisi. Le arterie ipofisarie superiori si dirigono verso il nucleo grigio e l'infundibolo dell'ipotalamo, anastomizzandosi tra loro e formando capillari che penetrano nel tessuto cerebrale: la rete emocapillare primaria. Dalle anse lunghe e corte di questa rete si formano le vene portali, che si dirigono verso il lobo anteriore dell'ipofisi. Nel parenchima del lobo anteriore dell'ipofisi, queste vene si dividono in ampi capillari sinusoidali, formando la rete emocapillare secondaria. Il lobo posteriore dell'ipofisi è irrorato principalmente dall'arteria ipofisaria inferiore. Tra le arterie ipofisarie superiore e inferiore si formano lunghe anastomosi arteriose. Il deflusso del sangue venoso dalla rete emocapillare secondaria avviene attraverso un sistema di vene che sfociano nei seni cavernosi e intercavernosi della dura madre del cervello.
Le fibre simpatiche che penetrano nell'organo insieme alle arterie partecipano all'innervazione dell'ipofisi. Le fibre nervose simpatiche postganglionari partono dal plesso dell'arteria carotide interna. Inoltre, numerose terminazioni dei processi delle cellule neurosecretorie situate nei nuclei dell'ipotalamo si trovano nel lobo posteriore dell'ipofisi.
Caratteristiche della ghiandola pituitaria legate all'età
Il peso medio dell'ipofisi nei neonati raggiunge 0,12 g. Il peso dell'organo raddoppia entro i 10 anni e triplica entro i 15. Entro i 20 anni, il peso dell'ipofisi raggiunge il suo massimo (530-560 mg) e rimane pressoché invariato nelle fasi successive. Dopo i 60 anni, si osserva una leggera diminuzione del peso di questa ghiandola endocrina.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Ormoni pituitari
L'unità della regolazione nervosa e ormonale nell'organismo è garantita dalla stretta connessione anatomica e funzionale tra l'ipofisi e l'ipotalamo. Questo complesso determina lo stato e il funzionamento dell'intero sistema endocrino.
La principale ghiandola endocrina che produce numerosi ormoni peptidici che regolano direttamente la funzione delle ghiandole periferiche è l'ipofisi. È una formazione di colore grigio-rossastro a forma di fagiolo, ricoperta da una capsula fibrosa del peso di 0,5-0,6 g. Varia leggermente a seconda del sesso e dell'età di una persona. Rimane la divisione generalmente accettata dell'ipofisi in due lobi che differiscono per sviluppo, struttura e funzione: quello anteriore distale - adenoipofisi - e quello posteriore - neuroipofisi. Il primo costituisce circa il 70% della massa totale della ghiandola ed è convenzionalmente suddiviso in parte distale, infundibolare e intermedia, il secondo in parte posteriore, o lobo, e peduncolo ipofisario. La ghiandola è situata nella fossa ipofisaria della sella turcica dell'osso sfenoide ed è collegata all'encefalo tramite il peduncolo. La parte superiore del lobo anteriore è ricoperta dal chiasma ottico e dai tratti ottici. L'afflusso di sangue all'ipofisi è molto abbondante e avviene tramite rami dell'arteria carotide interna (arterie ipofisarie superiore e inferiore), nonché tramite rami del circolo arterioso del cervello. Le arterie ipofisarie superiori partecipano all'afflusso di sangue all'adenoipofisi, mentre quelle inferiori alla neuroipofisi, entrando in contatto con le terminazioni neurosecretorie degli assoni dei nuclei a grandi cellule dell'ipotalamo. Le prime entrano nell'eminenza mediana dell'ipotalamo, dove si disperdono in una rete capillare (il plesso capillare primario). Questi capillari (con cui entrano in contatto i terminali degli assoni delle piccole cellule neurosecretorie dell'ipotalamo mediobasale) si raccolgono nelle vene portali che scendono lungo il peduncolo ipofisario nel parenchima dell'adenoipofisi, dove si dividono nuovamente in una rete di capillari sinusoidali (il plesso capillare secondario). Pertanto, il sangue, dopo aver attraversato l'eminenza mediana dell'ipotalamo, dove si arricchisce di ormoni adenoipofisotropici ipotalamici (ormoni di rilascio), entra nell'adenoipofisi.
Il deflusso di sangue saturo di ormoni adenoipofisari dai numerosi capillari del plesso secondario avviene attraverso il sistema venoso, che a sua volta confluisce nei seni venosi della dura madre e quindi nel flusso sanguigno generale. Pertanto, il sistema portale dell'ipofisi, con flusso sanguigno discendente dall'ipotalamo, è una componente morfofunzionale del complesso meccanismo di controllo neuroumorale delle funzioni trofiche dell'adenoipofisi.
L'ipofisi è innervata da fibre simpatiche che seguono le arterie ipofisarie. Originano da fibre postganglionari che attraversano il plesso carotideo interno, connesse ai gangli cervicali superiori. L'adenoipofisi non è innervata direttamente dall'ipotalamo. Il lobo posteriore riceve fibre nervose dai nuclei neurosecretori dell'ipotalamo.
L'adenoipofisi è una formazione molto complessa nella sua architettura istologica. Presenta due tipi di cellule ghiandolari: cromofobe e cromofile. Queste ultime si dividono a loro volta in acidofile e basofile (una descrizione istologica dettagliata dell'ipofisi è fornita nella sezione corrispondente del manuale). Tuttavia, è opportuno notare che gli ormoni prodotti dalle cellule ghiandolari che compongono il parenchima dell'adenoipofisi, a causa della diversità di quest'ultima, presentano una certa diversità chimica e la struttura fine delle cellule secernenti deve corrispondere alle caratteristiche biosintetiche di ciascuna di esse. Talvolta, tuttavia, nell'adenoipofisi si possono osservare forme transitorie di cellule ghiandolari in grado di produrre diversi ormoni. Esistono prove che il tipo di cellule ghiandolari dell'adenoipofisi non sia sempre determinato geneticamente.
Sotto il diaframma della sella turcica si trova la parte imbutiforme del lobo anteriore. Abbraccia il peduncolo dell'ipofisi, entrando in contatto con il tubercolo grigio. Questa parte dell'adenoipofisi è caratterizzata dalla presenza di cellule epiteliali e da un'abbondante vascolarizzazione. È anche ormonalmente attiva.
La parte intermedia (media) della ghiandola pituitaria è costituita da diversi strati di grandi cellule basofile secretorie-attive.
L'ipofisi svolge diverse funzioni attraverso i suoi ormoni. Il suo lobo anteriore produce ormoni adrenocorticotropi (ACTH), tireostimolanti (TSH), follicolostimolanti (FSH), luteinizzante (LH), lipotropici, nonché l'ormone della crescita somatotropo (STO) e la prolattina. Nel lobo intermedio viene sintetizzato l'ormone melanocita-stimolante (MSH), mentre nel lobo posteriore si accumulano vasopressina e ossitocina.
ACTH
Gli ormoni ipofisari sono un gruppo di ormoni proteici, peptidici e glicoproteine. Tra gli ormoni dell'ipofisi anteriore, l'ACTH è stato studiato meglio. È prodotto da cellule basofile. La sua principale funzione fisiologica è quella di stimolare la biosintesi e la secrezione di ormoni steroidei da parte della corteccia surrenale. L'ACTH mostra anche attività melanocitaria e lipotropica. Nel 1953 è stato isolato in forma pura. Successivamente, è stata stabilita la sua struttura chimica, costituita da 39 residui amminoacidici negli esseri umani e in diversi mammiferi. L'ACTH non ha specificità di specie. Attualmente, è stata effettuata la sintesi chimica sia dell'ormone stesso che di vari frammenti della sua molecola, più attivi degli ormoni naturali. La struttura dell'ormone presenta due sezioni della catena peptidica, una delle quali garantisce il rilevamento e il legame dell'ACTH al recettore, e l'altra fornisce un effetto biologico. Apparentemente, si lega al recettore dell'ACTH attraverso l'interazione delle cariche elettriche dell'ormone e del recettore. Il ruolo dell'effettore biologico dell'ACTH è svolto dal frammento della molecola 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
L'attività melanocitaria dell'ACTH è dovuta alla presenza nella molecola della regione N-terminale, composta da 13 residui amminoacidici e che ripete la struttura dell'ormone alfa-melanocita-stimolante. Questa regione contiene anche un eptapeptide presente in altri ormoni ipofisari e dotato di attività adrenocorticotropa, melanocitaria e lipotropica.
Il momento chiave nell'azione dell'ACTH è considerato l'attivazione dell'enzima protein chinasi nel citoplasma con la partecipazione dell'AMPc. La protein chinasi fosforilata attiva l'enzima esterasi, che converte gli esteri del colesterolo in una sostanza libera nelle goccioline lipidiche. La proteina sintetizzata nel citoplasma a seguito della fosforilazione dei ribosomi stimola il legame del colesterolo libero al citocromo P-450 e il suo trasferimento dalle goccioline lipidiche ai mitocondri, dove sono presenti tutti gli enzimi che assicurano la conversione del colesterolo in corticosteroidi.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Ormone stimolante la tiroide
Il TSH (tireotropina) è il principale regolatore dello sviluppo e del funzionamento della tiroide, nonché dei processi di sintesi e secrezione degli ormoni tiroidei. Questa proteina complessa, la glicoproteina, è composta da subunità alfa e beta. La struttura della prima subunità coincide con quella della subunità alfa dell'ormone luteinizzante. Inoltre, coincide ampiamente nelle diverse specie animali. La sequenza dei residui amminoacidici nella subunità beta del TSH umano è stata decifrata ed è composta da 119 residui amminoacidici. Si può notare che le subunità beta del TSH umano e bovino sono simili sotto molti aspetti. Le proprietà biologiche e la natura dell'attività biologica degli ormoni glicoproteici sono determinate dalla subunità beta. Essa garantisce inoltre l'interazione dell'ormone con i recettori in vari organi bersaglio. Tuttavia, nella maggior parte degli animali, la subunità beta mostra un'attività specifica solo dopo essersi combinata con la subunità alfa, che agisce come una sorta di attivatore dell'ormone. Quest'ultima, con altrettanta probabilità, induce attività luteinizzante, follicolo-stimolante e tireotropa, determinate dalle proprietà della subunità beta. La similarità riscontrata ci permette di concludere che questi ormoni siano derivati nel processo evolutivo da un predecessore comune; la subunità beta determina anche le proprietà immunologiche degli ormoni. Si presume che la subunità alfa protegga la subunità beta dall'azione degli enzimi proteolitici e ne faciliti il trasporto dall'ipofisi agli organi bersaglio periferici.
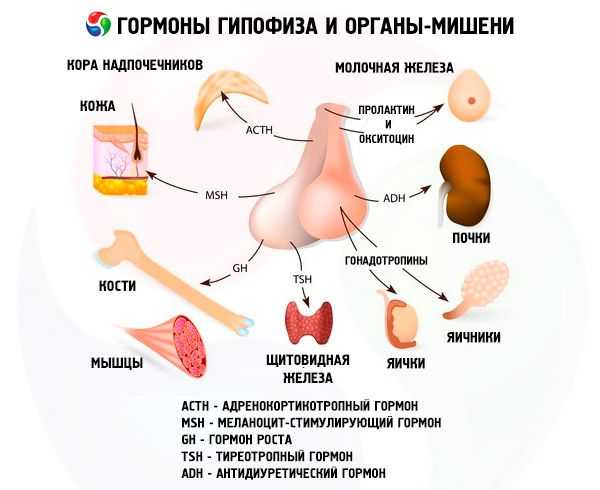
Ormoni gonadotropici
Le gonadotropine sono presenti nell'organismo sotto forma di LH e FSH. La funzione di questi ormoni è generalmente limitata a garantire i processi riproduttivi in individui di entrambi i sessi. Come il TSH, sono proteine complesse, le glicoproteine. L'FSH induce la maturazione dei follicoli nelle ovaie nelle donne e stimola la spermatogenesi negli uomini. L'LH causa la rottura del follicolo nelle donne con la formazione del corpo luteo e stimola la secrezione di estrogeni e progesterone. Negli uomini, lo stesso ormone accelera lo sviluppo del tessuto interstiziale e la secrezione di androgeni. Gli effetti delle gonadotropine sono interdipendenti e si verificano in modo sincrono.
La dinamica della secrezione di gonadotropine nelle donne cambia durante il ciclo mestruale ed è stata studiata in modo sufficientemente dettagliato. Nella fase preovulatoria (follicolare) del ciclo, il contenuto di LH è piuttosto basso, mentre quello di FSH è aumentato. Con la maturazione del follicolo, aumenta la secrezione di estradiolo, che favorisce una maggiore produzione di gonadotropine da parte dell'ipofisi e il verificarsi di cicli di LH e FSH, ovvero gli steroidi sessuali stimolano la secrezione di gonadotropine.
Attualmente, la struttura dell'LH è stata determinata. Come il TSH, è costituito da due subunità: a e b. La struttura della subunità alfa dell'LH nelle diverse specie animali coincide ampiamente, corrispondendo alla struttura della subunità alfa del TSH.
La struttura della subunità beta dell'LH differisce significativamente da quella della subunità beta del TSH, sebbene presenti quattro sezioni identiche della catena peptidica, composte da 4-5 residui amminoacidici. Nel TSH, sono localizzate nelle posizioni 27-31, 51-54, 65-68 e 78-83. Poiché la subunità beta di LH e TSH determina l'attività biologica specifica degli ormoni, si può supporre che sezioni omologhe nella struttura di LH e TSH garantiscano la connessione delle subunità beta con la subunità alfa, e che le sezioni con struttura diversa siano responsabili della specificità dell'attività biologica degli ormoni.
L'LH nativo è molto stabile all'azione degli enzimi proteolitici, ma la subunità beta viene rapidamente scissa dalla chimotripsina, mentre la subunità a è difficile da idrolizzare da parte dell'enzima, ovvero svolge una funzione protettiva, impedendo alla chimotripsina di accedere ai legami peptidici.
Per quanto riguarda la struttura chimica dell'FSH, i ricercatori non hanno ancora ottenuto risultati definitivi. Come l'LH, l'FSH è costituito da due subunità, ma la subunità β dell'FSH differisce da quella dell'LH.
Prolattina
Un altro ormone, la prolattina (ormone lattogenico), svolge un ruolo attivo nei processi riproduttivi. Le principali proprietà fisiologiche della prolattina nei mammiferi si manifestano sotto forma di stimolazione dello sviluppo delle ghiandole mammarie e dell'allattamento, della crescita delle ghiandole sebacee e degli organi interni. Promuove la manifestazione dell'effetto degli steroidi sui caratteri sessuali secondari nei maschi, stimola l'attività secretoria del corpo luteo nei topi e nei ratti e partecipa alla regolazione del metabolismo dei grassi. Negli ultimi anni, la prolattina ha ricevuto molta attenzione come regolatore del comportamento materno; tale polifunzionalità è spiegata dal suo sviluppo evolutivo. È uno degli ormoni ipofisari più antichi e si trova persino negli anfibi. Attualmente, la struttura della prolattina in alcune specie di mammiferi è stata completamente decifrata. Tuttavia, fino a poco tempo fa, gli scienziati nutrivano dubbi sull'esistenza di tale ormone negli esseri umani. Molti credevano che la sua funzione fosse svolta dall'ormone della crescita. Ora sono state ottenute prove convincenti della presenza di prolattina negli esseri umani e la sua struttura è stata parzialmente decifrata. I recettori della prolattina legano attivamente l'ormone della crescita e il lattogeno placentare, indicando un singolo meccanismo d'azione dei tre ormoni.
Somatotropina
L'ormone della crescita, la somatotropina, ha uno spettro d'azione ancora più ampio della prolattina. Come la prolattina, è prodotto dalle cellule acidofile dell'adenoipofisi. L'STH stimola la crescita scheletrica, attiva la biosintesi proteica, ha un effetto di mobilizzazione dei grassi e favorisce l'aumento delle dimensioni corporee. Inoltre, coordina i processi metabolici.
Il coinvolgimento dell'ormone in quest'ultimo è confermato dal fatto di un forte aumento della sua secrezione da parte della ghiandola pituitaria, ad esempio quando diminuisce il livello di zucchero nel sangue.
La struttura chimica di questo ormone umano è ormai completamente definita: 191 residui amminoacidici. La sua struttura primaria è simile a quella della somatomammotropina corionica o del lattogeno placentare. Questi dati indicano una significativa prossimità evolutiva tra i due ormoni, sebbene presentino differenze nell'attività biologica.
È necessario sottolineare l'elevata specificità di specie dell'ormone in questione: ad esempio, l'STH di origine animale è inattivo nell'uomo. Ciò è spiegato sia dalla reazione tra i recettori dell'STH di esseri umani e animali, sia dalla struttura dell'ormone stesso. Attualmente sono in corso studi per identificare i centri attivi nella complessa struttura dell'STH che mostrano attività biologica. Vengono studiati singoli frammenti della molecola che mostrano altre proprietà. Ad esempio, dopo l'idrolisi dell'STH umano da parte della pepsina, è stato isolato un peptide costituito da 14 residui amminoacidici e corrispondente alla sezione molecolare 31-44. Non ha avuto un effetto sulla crescita, ma ha superato significativamente l'ormone nativo in attività lipotropica. L'ormone della crescita umano, a differenza dell'ormone simile presente negli animali, ha una significativa attività lattogenica.
L'adenoipofisi sintetizza numerose sostanze peptidiche e proteiche che hanno un effetto di mobilizzazione dei grassi, e gli ormoni ipofisari trofici – ACTH, STH, TSH e altri – hanno un effetto lipotropico. Negli ultimi anni, gli ormoni beta- e γ-lipotropici (LPG) sono stati particolarmente enfatizzati. Le proprietà biologiche del beta-LPG sono state studiate in modo approfondito; oltre all'attività lipotropica, ha anche un effetto melanocita-stimolante, corticotropo-stimolante e ipocalcemico, e produce anche un effetto insulino-simile.
Attualmente, la struttura primaria dell'LPG ovino (90 residui amminoacidici) e degli ormoni lipotropici di suini e bovini è stata decifrata. Questo ormone è specie-specifico, sebbene la struttura della regione centrale del γ-LPG sia la stessa in specie diverse. Essa determina le proprietà biologiche dell'ormone. Uno dei frammenti di questa regione si trova nella struttura di γ-MSH, γ-MSH, ACTH e γ-LPG. Si suggerisce che questi ormoni derivino dallo stesso precursore nel processo evolutivo. Il γ-LPG ha un'attività lipotropica più debole del γ-LPG.
Ormone stimolante i melanociti
Questo ormone, sintetizzato nel lobo intermedio dell'ipofisi, stimola la biosintesi della melanina, il pigmento cutaneo, nella sua funzione biologica, e promuove l'aumento delle dimensioni e del numero di cellule pigmentate (melanociti) nella pelle degli anfibi. Queste proprietà dell'MSH sono utilizzate nei test biologici dell'ormone. Esistono due tipi di ormone: alfa-MSH e beta-MSH. È stato dimostrato che l'alfa-MSH non ha specificità di specie e ha la stessa struttura chimica in tutti i mammiferi. La sua molecola è una catena peptidica composta da 13 residui amminoacidici. Il beta-MSH, al contrario, ha specificità di specie e la sua struttura differisce nei diversi animali. Nella maggior parte dei mammiferi, la molecola del beta-MSH è composta da 18 residui amminoacidici e solo nell'uomo è estesa dall'estremità amminoacidica di quattro residui amminoacidici. È importante notare che l'alfa-MSH ha una certa attività adrenocorticotropa e il suo effetto sul comportamento di animali ed esseri umani è stato ormai dimostrato.
Ossitocina e vasopressina
La vasopressina e l'ossitocina, sintetizzate nell'ipotalamo, si accumulano nel lobo posteriore dell'ipofisi: la vasopressina nei neuroni del nucleo sopraottico e l'ossitocina nel nucleo paraventricolare. Successivamente, vengono trasferite all'ipofisi. È importante sottolineare che il precursore dell'ormone vasopressina viene sintetizzato per primo nell'ipotalamo. Contemporaneamente, vengono prodotte le proteine neurofisine di tipo 1 e 2. La prima lega l'ossitocina, la seconda la vasopressina. Questi complessi migrano sotto forma di granuli neurosecretori nel citoplasma lungo l'assone e raggiungono il lobo posteriore dell'ipofisi, dove le fibre nervose terminano nella parete vascolare e il contenuto dei granuli entra nel sangue. La vasopressina e l'ossitocina sono i primi ormoni ipofisari con una sequenza amminoacidica completamente definita. Nella loro struttura chimica sono nonapeptidi con un ponte disolfuro.
Gli ormoni in esame producono diversi effetti biologici: stimolano il trasporto di acqua e sali attraverso le membrane, hanno un effetto vasopressorio, potenziano le contrazioni della muscolatura liscia dell'utero durante il parto e aumentano la secrezione delle ghiandole mammarie. È importante notare che la vasopressina ha un'attività antidiuretica maggiore dell'ossitocina, mentre quest'ultima ha un effetto più forte sull'utero e sulla ghiandola mammaria. Il principale regolatore della secrezione di vasopressina è il consumo di acqua; nei tubuli renali, si lega ai recettori presenti sulle membrane citoplasmatiche con successiva attivazione dell'enzima adenilato ciclasi in esse presente. Diverse porzioni della molecola sono responsabili del legame dell'ormone al recettore e dell'effetto biologico.
L'ipofisi, collegata tramite l'ipotalamo a tutto il sistema nervoso, unisce il sistema endocrino in un insieme funzionale, contribuendo a garantire la costanza dell'ambiente interno dell'organismo (omeostasi). All'interno del sistema endocrino, la regolazione omeostatica avviene sulla base del principio di feedback tra il lobo anteriore dell'ipofisi e le ghiandole "bersaglio" (tiroide, corteccia surrenale, gonadi). Un eccesso dell'ormone prodotto dalla ghiandola "bersaglio" inibisce, mentre una sua carenza stimola la secrezione e il rilascio del corrispondente ormone trofico. L'ipotalamo è incluso nel sistema di feedback. È in esso che si trovano le zone recettoriali sensibili agli ormoni delle ghiandole "bersaglio". Legandosi specificamente agli ormoni circolanti nel sangue e modificando la risposta a seconda della concentrazione ormonale, i recettori ipotalamici trasmettono il loro effetto ai corrispondenti centri ipotalamici, che coordinano il lavoro dell'adenoipofisi, rilasciando ormoni adenoipofisiotropi ipotalamici. Pertanto, l'ipotalamo dovrebbe essere considerato un cervello neuroendocrino.