Esperto medico dell'articolo

Nuove pubblicazioni
Sintesi, secrezione e metabolismo delle catecolamine
Ultima recensione: 19.10.2021

Tutti i contenuti di iLive sono revisionati o verificati da un punto di vista medico per garantire la massima precisione possibile.
Abbiamo linee guida rigorose in materia di sourcing e colleghiamo solo a siti di media affidabili, istituti di ricerca accademici e, ove possibile, studi rivisti dal punto di vista medico. Nota che i numeri tra parentesi ([1], [2], ecc.) Sono link cliccabili per questi studi.
Se ritieni che uno qualsiasi dei nostri contenuti sia impreciso, scaduto o comunque discutibile, selezionalo e premi Ctrl + Invio.
Lo strato cerebrale delle ghiandole surrenali produce un composto lontano dagli steroidi della struttura. Contengono un nucleo di 3,4-diidrossifenile (catecolo) e sono chiamate catecolamine. Questi includono adrenalina, norepinefrina e dopamina beta-ossitiramina.
La sintesi delle catecolamine sequenza è abbastanza semplice: tirosina → diidrossifenilalanina (DOPA) → → → dopamina norepinefrina adrenalina. La tirosina entra nel corpo con il cibo, ma può anche essere formata dalla fenilalanina nel fegato sotto l'azione della fenilalanina idrossilasi. I prodotti finali della trasformazione della tirosina nei tessuti sono diversi. Nel midollo surrenale il processo procede al passo formazione adrenalina, alle estremità dei nervi simpatici - noradrenalina, in alcuni neuroni del sistema nervoso catecolamine sintesi della dopamina modulo compilato centrale.
La conversione della tirosina in DOPA è catalizzata dalla tirosina idrossilasi, i cui cofattori sono tetraidro-biopterina e ossigeno. Si ritiene che sia questo enzima che limita la velocità dell'intero processo della biosintesi delle catecolamine ed è inibito dai prodotti finali del processo. La tirosina idrossilasi è l'oggetto principale degli effetti regolatori sulla biosintesi delle catecolamine.
Dopa a dopamina conversione è catalizzata dall'enzima dopa-decarbossilasi (cofattore - pyridoxal) che è relativamente non specifici e decarbossilata, e altro acido aromatico L-ammino. Tuttavia, ci sono indicazioni sulla possibilità di modificare la sintesi delle catecolamine modificando l'attività e questo enzima. In alcuni neuroni non ci sono enzimi per l'ulteriore conversione della dopamina, ed è il prodotto finale. Altri tessuti contengono dopamina-beta-idrossilasi (i cofattori sono rame, acido ascorbico e ossigeno), che converte la dopamina in noradrenalina. Nella midollare surrenale (ma non alla fine dei nervi simpatici) c'è la feniletanolamina - metiltransferasi, che genera adrenalina dalla norepinefrina. Il donatore dei gruppi metilici in questo caso è l'S-adenosilmetionina.
È importante ricordare che la sintesi di Feniletanolammina-N-Metiltransferazy indotta da glucocorticoidi che rientrano nel cerebrale strato corticale del sistema venoso portale. In questo, forse, sta la spiegazione del fatto di combinare due diverse ghiandole endocrine in un organo. Sensi sintesi glucocorticoidi di adrenalina sottolineata dal fatto che le cellule midollo surrenale produrre noradrenalina, disposte attorno ai vasi arteriosi, mentre le cellule del sangue sono ottenuti adrenalinprodutsiruyuschie essenzialmente dei seni venosi, localizzata nella corteccia surrenale.
Il collasso delle catecolamine avviene principalmente sotto l'influenza dei due sistemi enzimatici: catecolo-O-metiltransferasi (COMT) e monoamino ossidasi (MAO). Le principali vie del decadimento di adrenalina e noradrenalina sono mostrate schematicamente in fig. 54. Sotto l'azione della COMT in presenza di un donatore di catecolamine gruppi metile S-adrenozilmetionina e convertito in normetanefrina metanefrina (3-O-metil-derivati di adrenalina e noradrenalina), che sotto l'influenza di MAO trasformati in aldeidi e più (in presenza di aldeide) in vanillil-mandelico acido (IUD) - il prodotto principale della disintegrazione della noradrenalina e dell'epinefrina. Nello stesso caso, quando prima esposto all'azione catecolamine MAO, non COMT, essi vengono convertiti in aldeide 3,4-dioksimindalevy, e poi sotto l'influenza della aldeide e COMT - acido 3,4-dioksimindalnuyu e la spirale. In presenza di alcol deidrogenasi di catecolamine può formare 3-metossi-4-oksifenilglikol, il principale prodotto finale della degradazione di adrenalina e noradrenalina nel SNC.
Disintegrazione dopamina procede in modo simile, eccetto che i suoi metaboliti sono privi di gruppi ossidrile sull'atomo di beta-carbonio, e quindi, invece di acido vanillil-mandelico omovanillico formata (HVA) e acido 3-metossi-4-oksifeniluksusnaya.
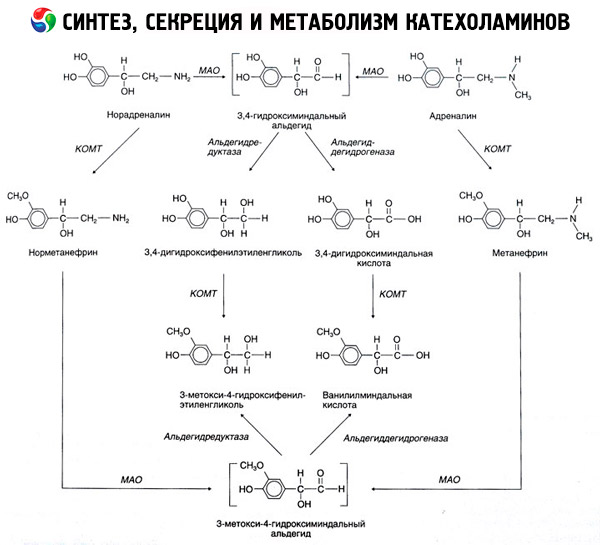
Si può anche postulare l'esistenza di un percorso quinoide per l'ossidazione della molecola delle catecolamine, su cui si ottengono prodotti intermedi con un'attività biologica pronunciata.
Formata dall'azione di enzimi citosolico, adrenalina e noradrenalina in terminazioni nervose simpatiche, midollare surrenale e inserire i granuli secretori che li protegge dall'azione degli enzimi degradativi. La cattura di catecolamine con granuli richiede costi energetici. In granuli cromaffini del midollo surrenale catecolamine saldamente legati all'ATP (nel rapporto di 4: 1) e le proteine specifiche - cromogranina che impedisce la diffusione degli ormoni dai granuli nel citoplasma.
Stimolo diretto alla secrezione di catecolamine è apparentemente penetrazione cella calcio stimolante esocitosi (granuli fusione di membrana con la superficie cellulare e la loro gap con il rendimento complessivo del titolo solubile - catecolamine, dopamina-beta-idrossilasi, ATP e cromogranina - nel fluido extracellulare) .
Effetti fisiologici delle catecolamine e del meccanismo della loro azione
Gli effetti delle catecolamine iniziano con l'interazione con specifici recettori delle cellule bersaglio. Se i recettori della tiroide e degli ormoni steroidei sono localizzati all'interno delle cellule, i recettori della catecolamina (così come l'acetilcolina e gli ormoni peptidici) sono presenti sulla superficie della cellula esterna.
Da tempo è stato stabilito che in relazione ad alcuni reazioni adrenalina o noradrenalina sono più efficaci rispetto alla isoproterenolo catecolamine sintetico, mentre per gli altri l'effetto è superiore all'azione di epinefrina isoproterenolo o noradrenalina. Su questa base, è stato sviluppato un concetto per la presenza nei tessuti di due tipi di adrenorecettori: alfa e beta, e in alcuni di essi può essere presente solo uno di questi due tipi. Isoproterenolo è il più potente agonista dei recettori beta-adrenergici, mentre fenilefrina composto sintetico - il più potente agonista dei recettori alfa-adrenergici. Catecolamine naturali - adrenalina e noradrenalina - sono in grado di interagire con i recettori di entrambi i tipi, ma l'adrenalina richiede una maggiore affinità per la beta, e noradrenalina - alfa-recettori.
Catecolamine più forti attivano recettori beta-adrenergici cardiaci di recettori beta del muscolo liscio, permettendo al beta-tipo suddivisi in sottotipi: beta1-recettori (cardiache, cellule di grasso) e recettori beta2 (bronchi, vasi sanguigni, ecc ...). L'azione di isoproterenolo sul recettore beta1 azione superiore di adrenalina e noradrenalina solo 10 volte, mentre i recettori beta2 agisce 100-1000 volte più potente di catecolamine naturale.
L'uso di antagonisti specifici (fentolamina e fenossibenzamina contro alfa e propranololo per i recettori beta) ha confermato l'adeguatezza della classificazione degli adrenorecettori. La dopamina è in grado di interagire con i recettori alfa e beta, ma in diversi tessuti (cervello, ipofisi, vasi), si trovano anche recettori dopaminergici il cui specifico bloccante è l'aloperidolo. Il numero di recettori beta varia da 1000 a 2000 per cellula. Gli effetti biologici delle catecolamine mediate dai recettori beta sono associati, di regola, all'attivazione di adenilato ciclasi e ad un aumento del contenuto intracellulare di cAMP. Il recettore e l'enzima, sebbene siano collegati funzionalmente, ma rappresentano macromolecole diverse. Nella modulazione dell'attività dell'adenilato ciclasi, sotto l'influenza del complesso ormone-recettore, sono coinvolti guanosina trifosfato (GTP) e altri nucleotidi purinici. Aumentando l'attività dell'enzima, sembrano ridurre l'affinità dei recettori beta per gli agonisti.
Il fenomeno di aumentare la sensibilità delle strutture denervate è noto da tempo. Viceversa, l'esposizione prolungata agli agonisti diminuisce la sensibilità dei tessuti bersaglio. Lo studio dei recettori beta ha permesso di spiegare questi fenomeni. È stato dimostrato che l'azione prolungata dell'isoproterenolo porta ad una perdita di sensibilità dell'adenilato ciclasi a causa di una diminuzione del numero di recettori beta.
Il processo di desensibilizzazione non richiede l'attivazione della sintesi proteica ed è probabilmente dovuto alla graduale formazione di complessi ormonali-recettori irreversibili. Al contrario, la somministrazione di 6-oxidofamin, che interrompe le estremità simpatiche, è accompagnata da un aumento del numero di recettori beta che reagiscono nei tessuti. Non è escluso che un aumento dell'attività nervosa simpatica determini la desensibilizzazione legata all'età dei vasi sanguigni e dei tessuti grassi in relazione alle catecolamine.
Il numero di adrenorecettori in diversi organi può essere controllato da altri ormoni. Pertanto, l'estradiolo aumenta e il progesterone riduce il numero di recettori alfa-adrenergici nell'utero, che è accompagnato da un corrispondente aumento e diminuzione nella sua risposta contrattile alle catecolamine. Se il "secondo messaggero" intracellulare formato dall'azione degli agonisti del recettore beta è certamente cAMP, allora il caso del trasmettitore di influenze alfa-adrenergiche è più complicato. Si presume che esistano vari meccanismi: una diminuzione del livello di cAMP, un aumento del contenuto di cAMP, una modulazione della dinamica cellulare del calcio,
Per riprodurre una varietà di effetti nel corpo, sono normalmente necessarie dosi di epinefrina, che sono 5-10 volte inferiori alla noradrenalina. Sebbene quest'ultimo sia più efficace per i recettori ad- e beta1-adrenergici, è importante ricordare che entrambe le catecolamine endogene sono in grado di interagire con i recettori alfa e beta. Pertanto, la risposta biologica di questo corpo all'attivazione adrenergica dipende in gran parte dal tipo di recettori presenti in esso. Tuttavia, questo non significa che l'attivazione selettiva del collegamento nervoso o umorale del sistema simpatico-surrenale sia impossibile. Nella maggior parte dei casi esiste un'intensa attività dei suoi vari collegamenti. Pertanto, si presume che esso attiva riflesso ipoglicemia midollare surrenale, mentre una diminuzione della pressione sanguigna (ipotensione posturale) accompagnata rilascio principalmente noradrenalina dalle terminazioni nervose simpatiche.
Adrenorecettori e gli effetti della loro attivazione in vari tessuti
|
Sistema, organo |
Tipo di adrenocettore |
Reazione |
|
Sistema cardiovascolare: |
||
|
Cuore |
Beta |
Aumento della frequenza delle contrazioni, della conduzione e della contrattilità |
|
Arteriole: |
||
|
Pelle e mucose |
Alfa |
Riduzione |
|
Dei muscoli scheletrici |
Beta |
Riduzione dell'estensione |
|
Organi addominali |
Alfa (altro) |
Riduzione |
|
Beta |
Estensione |
|
|
Vienna |
Alfa |
Riduzione |
|
Sistema respiratorio: |
||
|
Muscoli bronchiali |
Beta |
Estensione |
|
Il sistema digestivo: |
||
|
Stomaco |
Beta |
Diminuzione della funzione del motore |
|
Intestini |
Alfa |
Riduzione degli sfinteri |
|
Milza |
Alfa |
Riduzione |
|
Beta |
Relax |
|
|
Parte esternamente segreta del pancreas |
Alfa |
Diminuzione della secrezione |
|
Sistema genito-urinario: |
Alfa |
Riduzione dello sfintere |
|
Vescica |
Beta |
Rilassa il muscolo dell'esorcista |
|
Organi sessuali maschili |
Alfa |
Eiaculazione |
|
Occhi |
Alfa |
L'alunno si è dilatato |
|
Pelle |
Alfa |
Aumento della sudorazione |
|
Ghiandole salivari |
Alfa |
Isolamento di potassio e acqua |
|
Beta |
Secrezione di amilasi |
|
|
Ghiandole endocrine: |
||
|
Isolotti del pancreas |
||
|
Cellule beta |
Alfa (altro) |
Diminuzione della secrezione di insulina |
|
Beta |
Aumento della secrezione di insulina |
|
|
Cellule alfa |
Beta |
Aumento della secrezione di glucagone |
|
8 celle |
Beta |
Aumento della secrezione di somatostatina |
|
L'ipotalamo e l'ipofisi: |
||
|
Somatotrofы |
Alfa |
Aumento della secrezione di STH |
|
Beta |
Ridotta secrezione di STH |
|
|
Laktotrofы |
Alfa |
Diminuzione della secrezione di prolattina |
|
Tireotrofy |
Alfa |
Diminuzione della secrezione di TSH |
|
Kortikotrofy |
Alfa |
Aumento della secrezione di ACTH |
| beta | Diminuzione della secrezione di ACTH | |
|
Ghiandola tiroide: |
||
|
Cellule follicolari |
Alfa |
Diminuzione della secrezione di tiroxina |
|
Beta |
Aumento della secrezione di tiroxina |
|
|
Cellule parafollicolari (K) |
Beta |
Aumento della secrezione di calcitonina |
|
Le ghiandole paratiroidi |
Beta |
Aumento della secrezione di PTH |
|
Reni |
Beta |
Aumento della secrezione di renina |
|
Stomaco |
Beta |
Aumentare la secrezione di gastrina |
|
Scambio di base |
Beta |
Aumento del consumo di ossigeno |
|
Il fegato |
? |
Aumento della glicogenolisi e della gluconeogenesi dalla produzione di glucosio; aumentare la chetogenesi con il rilascio di corpi chetonici |
|
Tessuto adiposo |
Beta |
L'aumento della lipolisi con il rilascio di acidi grassi liberi e glicerolo |
|
Muscoli scheletrici |
Beta |
Aumento della glicolisi con il rilascio di piruvato e lattato; diminuzione della proteolisi con diminuzione della resa di alanina, glutammina |
E 'importante tenere a mente che i risultati della somministrazione endovenosa di catecolamine non sono sempre rispecchiano adeguatamente gli effetti dei composti endogeni. Ciò vale soprattutto per la noradrenalina, perché si trova in un futuro non corpo per lo più il sangue, ma direttamente nella fessura sinaptica. Pertanto noradrenalina endogena attiva, ad esempio, non solo i alfa recettori vascolari (aumento della pressione sanguigna), ma anche beta-recettori cardiaci (palpitazioni), mentre la somministrazione di fili esterni noradrenalina prevalentemente all'attivazione dei recettori alfa vascolare e riflesso (attraverso il vago) rallentamento battiti cardiaci.
Le basse dosi di epinefrina attivano principalmente i recettori beta dei vasi muscolari e del cuore, determinando un calo della resistenza vascolare periferica e un aumento del volume minuto del cuore. In alcuni casi, il primo effetto può predominare e, dopo la somministrazione di adrenalina, si sviluppa ipotensione. In dosi più elevate, l'adrenalina attiva anche i recettori alfa, che è accompagnata da un aumento della resistenza vascolare periferica e sullo sfondo di un aumento del volume minuto del cuore porta ad un aumento della pressione sanguigna. Tuttavia, il suo effetto sui recettori beta vascolari è anche conservato. Di conseguenza, l'aumento della pressione sistolica supera il valore simile della pressione diastolica (aumento della pressione del polso). Con l'introduzione di dosi ancora maggiori, gli effetti alfa-mimetici dell'epinefrina iniziano a prevalere: la pressione sistolica e diastolica aumenta in parallelo, sia sotto l'influenza della noradrenalina.
L'effetto delle catecolamine sul metabolismo è costituito dai loro effetti diretti e indiretti. I primi sono realizzati principalmente attraverso i beta-recettori. Processi più complessi sono associati al fegato. Sebbene il potenziamento della glicogenolisi epatica sia stato tradizionalmente considerato il risultato dell'attivazione del recettore beta, ci sono anche dati sul coinvolgimento dei recettori alfa in questo. Gli effetti mediati delle catecolamine sono associati alla modulazione della secrezione di molti altri ormoni, ad esempio l'insulina. Nell'azione dell'adrenalina sulla sua secrezione, la componente alfa-adrenergica predomina chiaramente, poiché è dimostrato che ogni stress è accompagnato dall'inibizione della secrezione di insulina.
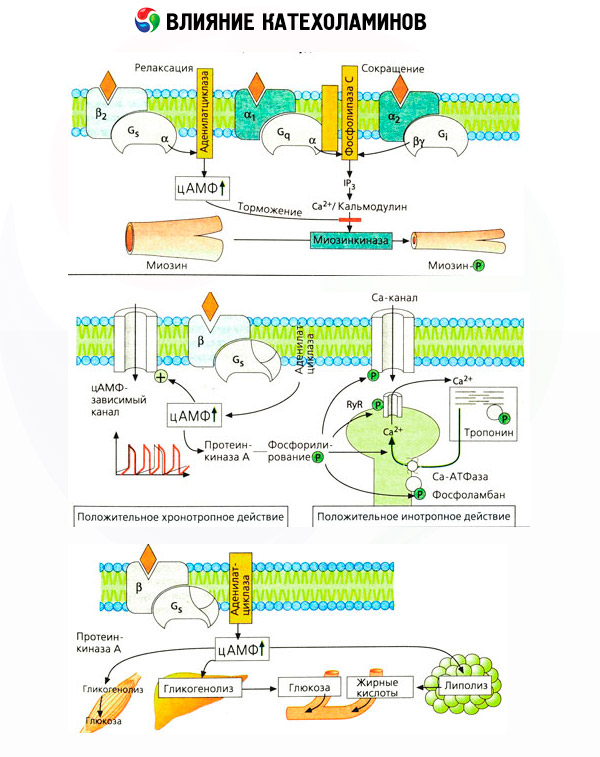
La combinazione di effetti diretti e indiretti delle catecolamine provoca iperglicemia, associata non solo ad un aumento della produzione epatica di glucosio, ma anche a inibizione del suo utilizzo da parte dei tessuti periferici. L'accelerazione della lipolisi causa l'iperlipidemia con aumento del rilascio di acidi grassi nel fegato e l'intensificazione della produzione di corpi chetonici. L'aumento della glicolisi nei muscoli porta ad un aumento del rilascio di lattato e piruvato nel sangue, che insieme al glicerolo rilasciato dal tessuto adiposo servono da precursori della gluconeogenesi epatica.
Regolazione della secrezione di catecolamine. La somiglianza dei prodotti e dei metodi di risposta del sistema nervoso simpatico e midollare surrenale è all'origine di combinare queste strutture in un unico corpo simpato neurale rilascio sistema ormonale e il suo collegamento. Vari segnali afferenti sono concentrati nell'ipotalamo e centri del midollo spinale e midollo allungato da cui emanano commutazione pacco efferente su corpi cellulari dei neuroni pregangliari situati nel corno laterale del midollo spinale a livello del VIII cervicale - segmenti lombari II-III.
Assoni pregangliari di queste cellule stanno lasciando il midollo spinale e formano connessioni sinaptiche con i neuroni sono localizzati nei gangli della catena simpatica, o le cellule midollo surrenale. Queste fibre pregangliari sono colinergiche. La prima differenza fondamentale di neuroni simpatici e postgangliari midollare surrenale cellule cromaffini consiste nel fatto che quest'ultimo viene trasmesso il segnale di ingresso è colinergici neuro-conduzione (postgangliari nervi adrenergici) ed umorale evidenziando composto adrenergici al sangue. La seconda differenza è ridotto a nervi postgangliari che producono noradrenalina, mentre le cellule midollo surrenale - preferibilmente adrenalina. Queste due sostanze hanno un effetto diverso sul tessuto.