Esperto medico dell'articolo

Nuove pubblicazioni
Clamidie
Ultima recensione: 06.07.2025

Tutti i contenuti di iLive sono revisionati o verificati da un punto di vista medico per garantire la massima precisione possibile.
Abbiamo linee guida rigorose in materia di sourcing e colleghiamo solo a siti di media affidabili, istituti di ricerca accademici e, ove possibile, studi rivisti dal punto di vista medico. Nota che i numeri tra parentesi ([1], [2], ecc.) Sono link cliccabili per questi studi.
Se ritieni che uno qualsiasi dei nostri contenuti sia impreciso, scaduto o comunque discutibile, selezionalo e premi Ctrl + Invio.
Le clamidie sono piccoli batteri parassiti coccoidi Gram-negativi appartenenti all'ordine Chlamydiales, famiglia Chlamydiaceae. Attualmente, questa famiglia comprende due generi che differiscono per struttura antigenica, inclusioni intracellulari e sensibilità ai sulfamidici: Chlamydia ( Chlamydia trachomatis ), Chlamydophila (Chlamydia pneumonia, Chlamydia psittaci ).
Il nome "clamidia" (dal greco chtamys, mantello) indica la presenza di una membrana che circonda le particelle microbiche.
Tutti i tipi di clamidia presentano caratteristiche morfologiche comuni, un antigene di gruppo comune e un ciclo riproduttivo distinto. Le clamidie sono considerate batteri Gram-negativi che hanno perso la capacità di sintetizzare ATP. Pertanto, sono parassiti energetici intracellulari obbligati.
Chlamydia trachomatis e Chlamydia pneumoniae sono classificate come microrganismi sicuramente patogeni per l'uomo e sono gli agenti causali della clamidia antropica. A seconda del tipo di agente patogeno e del punto di ingresso (vie respiratorie, apparato genitourinario), si distinguono clamidie respiratorie e urogenitali.
Sono state descritte più di 20 forme nosologiche causate da Chlamydia trachomatis, tra cui tracoma, congiuntivite, linfogranulomatosi inguinale, sindrome di Reiter e clamidia urogenitale. Le infezioni causate da Chlamydia trachomatis, secondo le stime dell'OMS, si collocano al secondo posto tra le malattie sessualmente trasmissibili dopo le infezioni da Trichomonas. Ogni anno si registrano circa 50 milioni di casi in tutto il mondo.
La polmonite da Chlamydophila causa una polmonite grave, una malattia delle vie respiratorie superiori. Si ipotizza che sia coinvolta nello sviluppo di aterosclerosi e asma bronchiale.
La Chlamydophila psittaci è la causa dell'ornitosi (psittacosi), una malattia zoonotica.
Proprietà morfologiche e tintoriali della clamidia
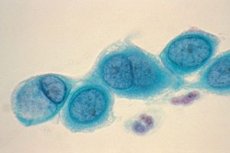
Le clamidie sono piccoli batteri Gram-negativi di forma sferica o ovoidale. Non hanno flagelli né capsule. Il metodo principale per rilevare la clamidia è la colorazione di Romanovsky-Giemsa. Il colore della colorazione dipende dallo stadio del ciclo vitale: gli anelli elementari sono colorati di viola su uno sfondo di citoplasma cellulare blu, i corpi reticolari sono colorati di blu.
La struttura della parete cellulare assomiglia a quella dei batteri Gram-negativi, sebbene vi siano delle differenze. Non contiene il tipico glicano peptidico: l'acido N-acetilmuramico è completamente assente dalla sua composizione. La parete cellulare include una membrana esterna, che include LPS e proteine. Nonostante l'assenza del glicano peptidico, la parete cellulare della clamidia è rigida. Il citoplasma cellulare è limitato da una membrana citoplasmatica interna.
L'analisi della membrana esterna (MO) della clamidia ha mostrato la presenza di LPS, la principale proteina della membrana esterna (MOMP), e delle proteine ricche di cisteina Ompl e Omp3, associate alla superficie interna della MOMP. LPS e MOMP di Chlamydia psittaci e Chlamydia trachomatis, a differenza delle MOMP di Chlamydia pneumoniae, sono localizzate sulla superficie esterna della cellula. Anche le proteine Omp di Chlamydia psittaci e Chlamydia pneumoniae, con un peso molecolare di 90-100 kDa, si trovano qui.
Le clamidie sono polimorfiche, il che è correlato alle peculiarità della loro riproduzione. Il ciclo di sviluppo unico (bifase) della clamidia è caratterizzato dall'alternanza di due diverse forme di esistenza: la forma infettiva (corpi elementari - EB) e la forma vegetativa (corpi reticolari, o iniziali - RT).
I microrganismi contengono RNA e DNA. Nella RT, l'RNA è 4 volte più abbondante del DNA. In questi, il contenuto è equivalente.
I corpi reticolari possono essere ovali, a forma di mezzaluna, sotto forma di bastoncelli bipolari e coccobacilli, di dimensioni comprese tra 300 e 1000 nm. I corpi reticolari non hanno proprietà infettive e, subendo la divisione, assicurano la riproduzione della clamidia.
Corpi elementari di forma ovale, di dimensioni comprese tra 250 e 500 nm, hanno proprietà infettive e sono in grado di penetrare in una cellula sensibile, dove avviene il ciclo di sviluppo. Hanno una membrana esterna densa, che li rende resistenti all'ambiente extracellulare.
Coltivazione della clamidia
Essendo parassiti obbligati, le clamidie non si riproducono su terreni nutritivi artificiali e possono essere coltivate solo in cellule viventi. Sono parassiti energetici, in quanto non sono in grado di accumulare energia autonomamente e di utilizzare l'ATP della cellula ospite. Le clamidie vengono coltivate in colture cellulari HeLa e McCoy, nel sacco vitellino di embrioni di pollo e nel corpo di animali sensibili a una temperatura di 35 °C.
 [ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
[ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
Struttura antigenica della clamidia
La clamidia ha tre tipi di antigeni: antigene specifico (comune a tutti i tipi di clamidia) - LPS; antigene info-specifico (diverso per tutti i tipi di clamidia) - di natura proteica, situato nella membrana esterna; tipo-specifico (diverso per i sierotipi di Chlamydia trachomatis) - LPS, che si moltiplica nella parete cellulare del microrganismo; antigene variante-specifico di natura proteica.
I sierotipi A, B e C sono detti oculari, in quanto causano il tracoma; i sierotipi D, E, K, O, H, I, J, K (geniale) sono gli agenti causali della clamidia urogenitale e delle sue complicanze; il sierotipo L è l'agente causale della linfogranulomatosi venerea. L'agente causale della clamidia respiratoria è Chlamydia pneumoniae, che ha 4 sierotipi: TWAR, AR, RF, CWL. Chlamydia psittaci ha 13 sierotipi.
[ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Tropismo cellulare della clamidia
La Chlamydia trachomatis ha un tropismo per la mucosa dell'epitelio del tratto urogenitale e può rimanere localmente su di essa o diffondersi su tutta la superficie del tessuto. L'agente eziologico del linfogranuloma venereo ha un tropismo per il tessuto linfoide.
La Chlamydia pneumoniae si replica nei macrofagi alveolari, nei monociti e nelle cellule endoteliali vascolari; è possibile anche la disseminazione sistemica dell'infezione.
La Chlamydia psittaci causa infezioni in vari tipi di cellule, compresi i fagociti mononucleari.
Ciclo vitale della clamidia
Il ciclo di sviluppo della clamidia dura 40-72 ore e comprende due forme di esistenza che differiscono per proprietà morfologiche e biologiche.
Nella prima fase del processo infettivo, l'adsorbimento dei corpi elementari della clamidia sulla membrana plasmatica della cellula ospite sensibile avviene grazie alla partecipazione di forze elettrostatiche. L'introduzione della clamidia nella cellula avviene per endocitosi. Le porzioni di membrana plasmatica su cui sono adsorbiti gli embrioni elementari si invaginano nel citoplasma con la formazione di vacuoli fagocitari. Questa fase dura 7-10 ore.
Successivamente, entro 6-8 ore, i corpi elementari infettivi si riorganizzano in forme intracellulari vegetative, metabolicamente attive e non infettive (RT), che si dividono ripetutamente. Queste forme intracellulari, che sono microcolonie, sono chiamate inclusioni clamidiali. Entro 18-24 ore dallo sviluppo, si localizzano in una vescicola citoplasmatica formata dalla membrana della cellula ospite. L'inclusione può contenere da 100 a 500 corpi reticolari di clamidia.
Nella fase successiva, durante le 36-42 ore, si verifica la maturazione (formazione di corpi intermedi) e la trasformazione dei corpi reticolari per divisione in corpi elementari. La cellula infetta viene distrutta e i corpi elementari la abbandonano. Essendo extracellulari, i corpi elementari penetrano in nuove cellule ospiti dopo 40-72 ore, dando inizio a un nuovo ciclo di sviluppo della clamidia.
Oltre a questo ciclo riproduttivo, in condizioni sfavorevoli si realizzano altri meccanismi di interazione tra la clamidia e la cellula ospite: la distruzione della clamidia nei fagosomi, la trasformazione L-like e la persistenza.
Le forme trasformate e persistenti della clamidia sono in grado di ritornare alle forme originali (reticolari) con successiva trasformazione in corpi elementari.
All'esterno delle cellule ospiti, le funzioni metaboliche sono ridotte al minimo.
Fattori di patogenicità della clamidia
Le proprietà adesive della clamidia sono dovute alle proteine della membrana esterna delle cellule, che possiedono anche proprietà antifagocitarie. Inoltre, le cellule microbiche contengono endotossine e producono esotossine. Le endotossine sono rappresentate da LPS, per molti aspetti simili a quelle dei batteri Gram-negativi. La termolabilità della sostanza è dovuta alle esotossine, presenti in tutti i batteri e che causano la morte dei topi dopo somministrazione endovenosa.
È stato scoperto che la clamidia ha un sistema secretorio di tipo III, attraverso il quale le proteine della clamidia vengono iniettate nel citoplasma della cellula ospite come parte integrante del processo infettivo.
La proteina da shock termico (HSP) ha la capacità di indurre reazioni autoimmuni.
[ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ], [ 34 ]
Ecologia e resistenza della clamidia
Le clamidie sono microrganismi molto comuni. Sono state trovate in oltre 200 specie di animali, pesci, anfibi, molluschi e artropodi. Microrganismi con morfologia simile sono stati trovati anche nelle piante superiori. I principali ospiti delle clamidie sono l'uomo, gli uccelli e i mammiferi.
L'agente eziologico della clamidia è instabile nell'ambiente esterno, è molto sensibile alle alte temperature e muore rapidamente una volta essiccato. La sua inattivazione a 50 °C avviene dopo 30 minuti, a 90 °C dopo 1 minuto. A temperatura ambiente (18-20 °C), l'attività infettiva del patogeno diminuisce dopo 5-7 giorni. A 37 °C, si osserva un calo della virulenza dell'80% dopo 6 ore in un termostato. Le basse temperature (-20 °C) contribuiscono alla conservazione a lungo termine delle proprietà infettive del patogeno. La clamidia muore rapidamente sotto l'influenza dei raggi UV, per contatto con etere etilico ed etanolo al 70%, sotto l'influenza di lisolo al 2% per 10 minuti, cloramina al 2%.