Esperto medico dell'articolo

Nuove pubblicazioni
Ipotalamo
Ultima recensione: 07.07.2025

Tutti i contenuti di iLive sono revisionati o verificati da un punto di vista medico per garantire la massima precisione possibile.
Abbiamo linee guida rigorose in materia di sourcing e colleghiamo solo a siti di media affidabili, istituti di ricerca accademici e, ove possibile, studi rivisti dal punto di vista medico. Nota che i numeri tra parentesi ([1], [2], ecc.) Sono link cliccabili per questi studi.
Se ritieni che uno qualsiasi dei nostri contenuti sia impreciso, scaduto o comunque discutibile, selezionalo e premi Ctrl + Invio.
L'ipotalamo forma le sezioni inferiori del diencefalo e partecipa alla formazione del pavimento del terzo ventricolo. L'ipotalamo comprende il chiasma ottico, il tratto ottico, il tubercolo grigio con l'imbuto e i corpi mammillari.
Il chiasma ottico (chiasma opticum) è una cresta trasversale formata dalle fibre dei nervi ottici (II paio di nervi cranici), che si incrociano parzialmente sul lato opposto (formando una decussazione). Questa cresta si continua lateralmente e posteriormente su ciascun lato nel tratto ottico (tratus opticus). Il tratto ottico si trova medialmente e posteriormente rispetto alla sostanza perforata anteriore, si piega attorno al peduncolo cerebrale dal lato laterale e termina con due radici nei centri visivi sottocorticali. La radice laterale, più grande (radix lateralis), si avvicina al corpo genicolato laterale, mentre la radice mediale, più sottile (radix medialis), si dirige verso il collicolo superiore del tetto del mesencefalo.
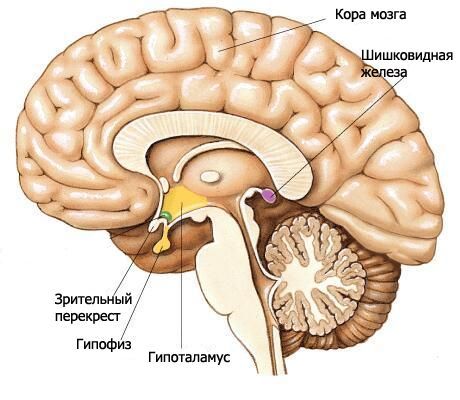
La placca terminale, che appartiene al telencefalo, è adiacente alla superficie anteriore del chiasma ottico e si fonde con esso. Chiude la sezione anteriore della scissura longitudinale del cervello ed è costituita da un sottile strato di sostanza grigia, che nelle sezioni laterali della placca si prolunga nella sostanza dei lobi frontali degli emisferi.
Dietro il chiasma ottico si trova il tubercolo grigio (tuber cinereum), dietro il quale si trovano i corpi mammillari, e ai lati si trovano i tratti ottici. Al di sotto, il tubercolo grigio passa nell'imbuto (infundibolo), che si collega all'ipofisi. Le pareti del tubercolo grigio sono formate da una sottile lamina di sostanza grigia contenente i nuclei grigio-tuberali (nuclei tuberales). Dal lato della cavità del terzo ventricolo, una depressione ristretta dell'imbuto si proietta nella regione del tubercolo grigio e ulteriormente all'interno dell'imbuto.
I corpi mammillari (corpora mamillaria) sono situati tra il tubercolo grigio anteriore e la sostanza perforata posteriore. Hanno l'aspetto di due piccole formazioni sferiche bianche, di circa 0,5 cm di diametro ciascuna. La sostanza bianca è situata solo all'esterno del corpo mammillare. All'interno si trova la sostanza grigia, in cui si distinguono i nuclei mediale e laterale del corpo mammillare (nuclei corporis mamillaris mediales et laterales). Le colonne del fornice terminano nei corpi mammillari.
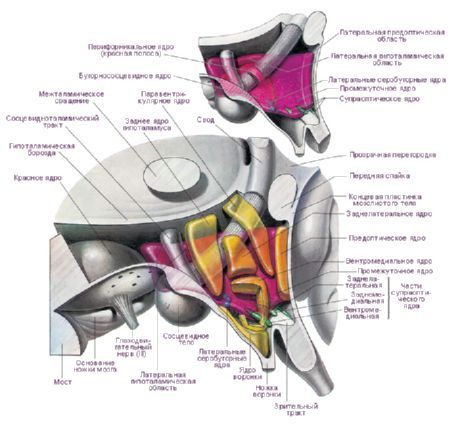
Nell'ipotalamo si trovano tre regioni ipotalamiche principali: gruppi di cellule nervose di diverse forme e dimensioni: anteriore (regio hypothalamica anteriore), intermedia (regio hypothalamica intermedia) e posteriore (regio hypothalamica posteriore). I gruppi di cellule nervose in queste regioni formano più di 30 nuclei dell'ipotalamo.
Le cellule nervose dei nuclei ipotalamici hanno la capacità di produrre una secrezione (neurosecrezione), che può essere trasportata all'ipofisi attraverso i processi di queste stesse cellule. Tali nuclei sono chiamati nuclei neurosecretori dell'ipotalamo. Nella regione anteriore dell'ipotalamo si trovano il nucleo sopraottico (nucleus supraopticus) e i nuclei paraventricolari (nuclei paraventriculares). I processi delle cellule di questi nuclei formano il fascio ipotalamo-ipofisario, che termina nel lobo posteriore dell'ipofisi. Tra il gruppo di nuclei della regione posteriore dell'ipotalamo, i più grandi sono i nuclei mediale e laterale del corpo mammillare (nuclei corporis mamillaris mediales et laterales) e il nucleo ipotalamico posteriore (nucleus hypothalamicus posterior). Il gruppo dei nuclei della regione ipotalamica intermedia comprende i nuclei ipotalamici inferomediali e superomediali (nuclei hypothalamic ventromediales et dorsomediales), il nucleo ipotalamico dorsale (nucleus hypothalamicus dorsalis), il nucleo dell'infundibolare (nucleus infundibularis), i nuclei grigio-tuberosi (nuclei tuberales), ecc.
I nuclei dell'ipotalamo sono collegati da un sistema piuttosto complesso di vie afferenti ed efferenti. Pertanto, l'ipotalamo ha un effetto regolatore su numerose funzioni vegetative dell'organismo. La neurosecrezione dei nuclei dell'ipotalamo può influenzare le funzioni delle cellule ghiandolari dell'ipofisi, aumentando o inibendo la secrezione di numerosi ormoni, che a loro volta regolano l'attività di altre ghiandole endocrine.
La presenza di connessioni neurali e umorali tra i nuclei ipotalamici e l'ipofisi ha reso possibile la loro fusione nel sistema ipotalamo-ipofisario.
Studi filogenetici hanno dimostrato che l'ipotalamo è presente in tutti i cordati, è ben sviluppato negli anfibi e ancor di più nei rettili e nei pesci. Gli uccelli presentano una differenziazione dei nuclei chiaramente espressa. Nei mammiferi, la materia grigia è altamente sviluppata, le cui cellule si differenziano in nuclei e campi. L'ipotalamo umano non differisce significativamente dall'ipotalamo dei mammiferi superiori.
Esistono numerose classificazioni dei nuclei ipotalamici. E. Gruntel ha identificato 15 coppie di nuclei, W. Le Gros Clark 16, H. Kuhlenbek 29. La classificazione più utilizzata è quella di W. Le Gros Clark. IN Bogolepova (1968), sulla base delle classificazioni sopra menzionate e tenendo conto dei dati ontogenetici, propone di suddividere i nuclei ipotalamici in quattro sezioni:
- sezione anteriore o rostrale (che combina l'area preottica e il gruppo anteriore - W. Le Gros Clark) - aree preottiche mediale e laterale, nucleo soprachiasmatico, nucleo sopraottico, nucleo paraventricolare, area ipotalamica anteriore;
- sezione mediale mediale - nucleo ventromediale, nucleo dorsomediale, nucleo infundibolare, area ipotalamica posteriore;
- sezione laterale media - area ipotalamica laterale, nucleo ipotalamico laterale, nucleo tuberolaterale, nucleo tuberomammillare, nucleo perifornicale;
- sezione posteriore o mammillare: nucleo mammillare mediale, nucleo mammillare laterale.
Le connessioni anatomiche dell'ipotalamo ne chiariscono anche il significato (funzionale). Tra le vie afferenti più importanti, si possono distinguere le seguenti:
- il fascio proencefalico mediale, la cui parte laterale collega l'ipotalamo con il bulbo olfattivo e il tubercolo, la regione periamigdaloidea e l'ippocampo, e la parte mediale con il setto, la regione diagonale e il nucleo caudato;
- la striscia terminale, che va dalla tonsilla alle parti anteriori dell'ipotalamo;
- fibre che passano attraverso il fornice dall'ippocampo al corpo mammillare;
- connessioni talamo-, strio- e pallidoipotalamiche;
- dal tronco encefalico - il tratto tegmentale centrale;
- dalla corteccia cerebrale (orbitale, temporale, parietale).
Pertanto, le principali fonti di afferentazione sono le formazioni limbiche del proencefalo e la formazione reticolare del tronco encefalico.
Anche i sistemi efferenti dell'ipotalamo possono essere raggruppati in tre direzioni:
- sistemi discendenti alla formazione reticolare e al midollo spinale: il sistema periventricolare di fibre che terminano nel mesencefalo (fascio longitudinale posteriore), nei centri autonomi del tronco caudale e del midollo spinale, e il fascio mammillare-tegmentale, che va dai corpi mammillari alla formazione reticolare del mesencefalo;
- percorsi verso il talamo dai corpi mammillari (fascio mammatalamico), che fanno parte del sistema limbico funzionale chiuso;
- Vie che portano all'ipofisi: la via ipotalamo-ipofisaria dai nuclei paraventricolare (10-20% delle fibre) e sopraottico (80-90%) ai lobi posteriori e parzialmente medi dell'ipofisi, la via tuberoipofisaria dai nuclei ventromediale e infundibolare all'adenoipofisi.
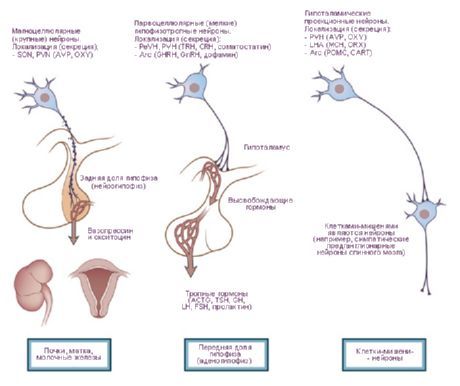
I lavori di J. Ranson (1935) e W. Hess (1930, 1954, 1968) presentarono dati sulla dilatazione e costrizione della pupilla, sull'aumento e la diminuzione della pressione arteriosa, sull'accelerazione e decelerazione del polso in seguito a stimolazione dell'ipotalamo. Sulla base di questi studi, furono identificate zone che esercitano effetti simpatici (sezione posteriore dell'ipotalamo) e parasimpatici (sezione anteriore), e l'ipotalamo stesso fu considerato un centro che integra l'attività del sistema viscerale che innerva organi e tessuti. Tuttavia, con lo sviluppo di questi studi, furono rivelati anche numerosi effetti somatici, soprattutto durante il comportamento libero degli animali [Gellhorn E., 1948]. O. G. Baklavadzhan (1969), in seguito a stimolazione di diverse sezioni dell'ipotalamo, osservò in alcuni casi una reazione di attivazione nella corteccia cerebrale, una facilitazione dei potenziali monosinaptici del midollo spinale, un aumento della pressione arteriosa e, in altri, l'effetto opposto. In questo caso, le reazioni vegetative presentavano la soglia più alta. O. Sager (1962) scoprì l'inibizione del sistema y e la sincronizzazione EEG durante la diatermia dell'ipotalamo, e l'effetto opposto durante un riscaldamento eccessivo. Si forma l'idea dell'ipotalamo come parte del cervello che svolge l'interazione tra meccanismi regolatori e l'integrazione dell'attività somatica e vegetativa. Da questo punto di vista, è più corretto dividere l'ipotalamo non in parti simpatiche e parasimpatiche, ma distinguerne le zone dinamogeniche (ergotropiche e trofotropiche). Questa classificazione è di natura funzionale e biologica e riflette il coinvolgimento dell'ipotalamo nell'attuazione di azioni comportamentali olistiche. Ovviamente, non solo il sistema vegetativo, ma anche quello somatico partecipa al mantenimento dell'omeostasi. Le zone ergotropiche e trofotropiche sono localizzate in tutte le parti dell'ipotalamo e si sovrappongono in alcune aree. Allo stesso tempo, è possibile identificare le zone di "concentrazione". Pertanto, nelle sezioni anteriori (zona preottica) gli apparati trofotropici sono rappresentati più chiaramente, mentre nelle sezioni posteriori (corpi mammillari) quelli ergotropici. L'analisi delle principali connessioni afferenti ed efferenti dell'ipotalamo con i sistemi limbico e reticolare fa luce sul suo ruolo nell'organizzazione di forme integrative di comportamento. L'ipotalamo occupa una posizione speciale, centrale, in questo sistema sia per la sua posizione topografica al centro di queste formazioni, sia per caratteristiche fisiologiche. Quest'ultima è determinata dal ruolo dell'ipotalamo come sezione del cervello specificamente costruita, particolarmente sensibile ai cambiamenti nell'ambiente interno del corpo, che reagisce alle più piccole fluttuazioni negli indicatori umorali e forma opportuni atti comportamentali in risposta a questi cambiamenti.Il ruolo specifico dell'ipotalamo è determinato dalla sua vicinanza anatomica e funzionale all'ipofisi. I nuclei dell'ipotalamo si dividono in specifici e aspecifici. Il primo gruppo comprende formazioni che si proiettano sull'ipofisi, il resto comprende altri nuclei, i cui effetti di stimolazione possono variare a seconda dell'intensità dell'impatto. I nuclei specifici dell'ipotalamo hanno un effetto evidente e si differenziano dalle altre formazioni cerebrali per la loro capacità di neurocrinia. Tra questi rientrano i nuclei sopraottico, paraventricolare e parvocellulare del tubercolo grigio. È stato accertato che l'ormone antidiuretico (ADH) viene prodotto nei nuclei sopraottico e paraventricolare, che discende lungo gli assoni del tratto ipotalamo-ipofisario fino al lobo posteriore dell'ipofisi. Successivamente è stato dimostrato che i fattori di rilascio si formano nei neuroni dell'ipotalamo e, entrando nell'adenoipofisi, regolano la secrezione di tre ormoni: adrenocorticotropo (ACTH), luteinizzante (LH), follicolo-stimolante (FSH) e tireostimolante (TSH). Le zone di formazione dei fattori di rilascio per ACTH e TSH sono i nuclei della parte anteriore dell'eminenza mediana e dell'area preottica, e per GTG le parti posteriori del tubercolo grigio. È stato stabilito che i fasci ipotalamo-ipofisari nell'uomo contengono circa 1 milione di fibre nervose.
Indubbiamente, anche altre parti del cervello (strutture medio-basali della regione temporale, formazione reticolare del tronco encefalico) partecipano alla regolazione neuroendocrina. Tuttavia, l'apparato più specifico è l'ipotalamo, che include le ghiandole endocrine nel sistema di reazioni integrali dell'organismo, in particolare le reazioni di natura stressante. I sistemi trofo- ed ergotropico dispongono non solo dei sistemi periferici simpatico e parasimpatico per garantire l'attività, ma anche di specifici apparati neuroormonali. Il sistema ipotalamo-ipofisario, che funziona secondo il principio del feedback, è in gran parte autoregolato. L'attività della formazione dei fattori di attivazione è determinata anche dal livello di ormoni nel sangue periferico.
L'ipotalamo è quindi una componente importante del sistema limbico e reticolare del cervello, ma essendo incluso in questi sistemi conserva i suoi specifici "input" sotto forma di speciale sensibilità ai cambiamenti dell'ambiente interno, nonché specifici "output" attraverso il sistema ipotalamo-ipofisario, connessioni paraventricolari con le formazioni vegetative sottostanti, nonché attraverso il talamo e la formazione reticolare del tronco encefalico verso la corteccia e il midollo spinale.
 [
[